

- Research Scientist and Consultant, Deerfield, NH 03037, USA
- Spoken Language Systems Group, Computer Science and Artificial Intelligence Laboratory, MIT, Cambridge MA 02139, USA
Correspondence Address:
Stephanie Seneff
Spoken Language Systems Group, Computer Science and Artificial Intelligence Laboratory, MIT, Cambridge MA 02139, USA
DOI:10.4103/2152-7806.153876
Copyright: © 2015 Samsel A. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.How to cite this article: Samsel A, Seneff S. Glyphosate, pathways to modern diseases III: Manganese, neurological diseases, and associated pathologies. Surg Neurol Int 24-Mar-2015;6:45
How to cite this URL: Samsel A, Seneff S. Glyphosate, pathways to modern diseases III: Manganese, neurological diseases, and associated pathologies. Surg Neurol Int 24-Mar-2015;6:45. Available from: http://surgicalneurologyint.com/surgicalint_articles/glyphosate-pathways-to-modern-diseases-iii-manganese-neurological-diseases-and-associated-pathologies/
Abstract
Manganese (Mn) is an often overlooked but important nutrient, required in small amounts for multiple essential functions in the body. A recent study on cows fed genetically modified Roundup®-Ready feed revealed a severe depletion of serum Mn. Glyphosate, the active ingredient in Roundup®, has also been shown to severely deplete Mn levels in plants. Here, we investigate the impact of Mn on physiology, and its association with gut dysbiosis as well as neuropathologies such as autism, Alzheimer's disease (AD), depression, anxiety syndrome, Parkinson's disease (PD), and prion diseases. Glutamate overexpression in the brain in association with autism, AD, and other neurological diseases can be explained by Mn deficiency. Mn superoxide dismutase protects mitochondria from oxidative damage, and mitochondrial dysfunction is a key feature of autism and Alzheimer’s. Chondroitin sulfate synthesis depends on Mn, and its deficiency leads to osteoporosis and osteomalacia. Lactobacillus, depleted in autism, depend critically on Mn for antioxidant protection. Lactobacillus probiotics can treat anxiety, which is a comorbidity of autism and chronic fatigue syndrome. Reduced gut Lactobacillus leads to overgrowth of the pathogen, Salmonella, which is resistant to glyphosate toxicity, and Mn plays a role here as well. Sperm motility depends on Mn, and this may partially explain increased rates of infertility and birth defects. We further reason that, under conditions of adequate Mn in the diet, glyphosate, through its disruption of bile acid homeostasis, ironically promotes toxic accumulation of Mn in the brainstem, leading to conditions such as PD and prion diseases.
Keywords: Autism, cholestasis, glyphosate, manganese, Parkinson's disease
INTRODUCTION
Glyphosate is the active ingredient in Roundup®, the most widely used herbicide on the planet.[
It may seem implausible that glyphosate could be toxic to humans, given the fact that government regulators appear nonchalant about steadily increasing residue limits, and that the levels in food and water are rarely monitored by government agencies, presumably due to lack of concern. However, a paper by Antoniou et al.[
Adjuvants in pesticides are synergistically toxic with the active ingredient. Mesnage et al.[
The industry dictates that 3 months is a sufficiently long time to test for toxicity in rodent studies, and as a consequence none of the industry studies have run for longer than 3 months. The only study we are aware of that was a realistic assessment of the long-term effects of GM Roundup®-Ready corn and soy feed on mammals was the study by Séralini et al. that examined the effects on rats fed these foods for their entire life span.[
There are multiple pathways by which glyphosate could lead to pathology.[
Metal chelation and inactivation of cytochrome P450 (CYP) enzymes (which contain heme) play important roles in the adverse effects of glyphosate on humans. A recent study on rats showed that both males and females exposed to Roundup® had 50% reduction in hepatic CYP enzyme levels compared with controls.[
Glyphosate is a likely cause of the recent epidemic in celiac disease.[
Glyphosate is also neurotoxic.[
In the remainder of this paper, we first introduce the link between glyphosate and manganese (Mn) dysbiosis, and briefly describe the main biological roles of Mn. We then describe how glyphosate's disruption of gut bacteria may be a major player in the recent epidemic in antibiotic resistance. We then explain how glyphosate can influence the uptake of arsenic and aluminum, and propose similar mechanisms at work with Mn. In the next section, we describe how Mn deficiency can lead to a reduction in Lactobacillus in the gut, and we link this to anxiety disorder. We follow with a discussion on mitochondrial dysfunction associated with suppressed Mn superoxide dismutase (Mn-SOD), and then a section on implications of Mn deficiency for oxalate metabolism. The following section explains how Mn deficiency can lead to the overexpression of ammonia and glutamate in many neurological diseases. The next two sections show how Mn accumulation in the liver is linked to cholestasis and high serum low density lipoprotein (LDL), and how this can also induce increased susceptibility to Salmonella poisoning. We then identify a role for Mn in chondroitin sulfate synthesis, and the implications for osteomalacia. The next two sections explain how glyphosate exposure can lead to Mn toxicity in the brain, and discuss two neurological diseases that are associated with excess Mn, PD and prion diseases. After a section on the link between male infertility and Mn deficiency in the testes, we discuss evidence of exposure to glyphosate and end with a short summary of our findings.
SUPPORTIVE EVIDENCE OF MANGANESE DYSBIOSIS DUE TO GLYPHOSATE
Glyphosate's disruption of the shikimate pathway is due in part to its chelation of Mn, which is a catalyst for enolpyruvylshikimate phosphate synthase (EPSPS), a critical early enzyme in the pathway.[
Mn, named after the Greek word for “magic,” is one of 14 essential trace elements. Mn plays essential roles in antioxidant protection, glutamine synthesis, bone development, and sperm motility, among other things. Although Mn is essential, it is only required in trace amounts. And an excess of Mn can be neurotoxic.
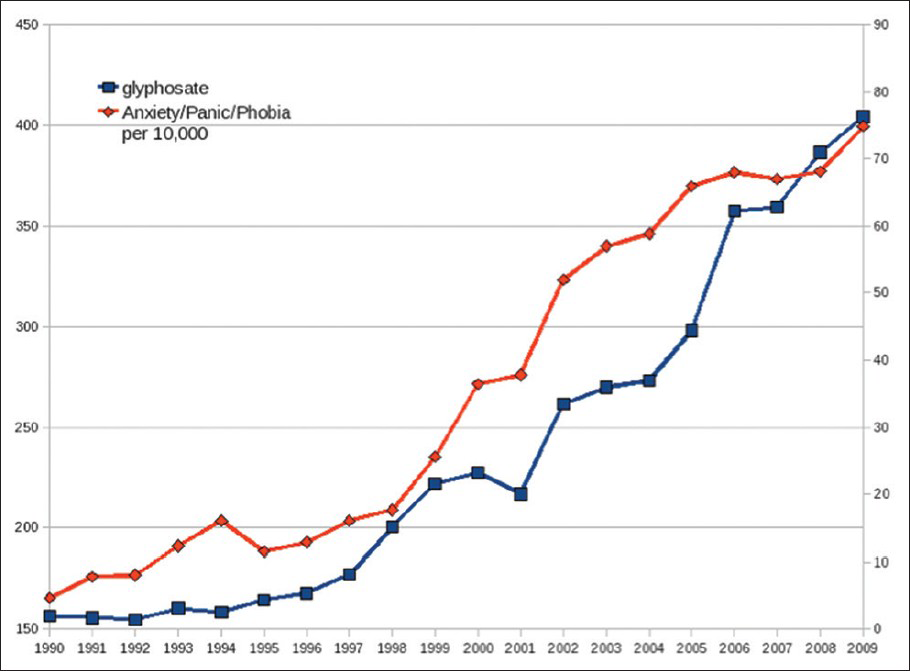
Remarkably, Mn deficiency can explain many of the pathologies associated with autism and Alzheimer's disease (AD). The incidence of both of these conditions has been increasing at an alarming rate in the past two decades, in step with the increased usage of glyphosate on corn and soy crops in the United States, as shown in [Figures
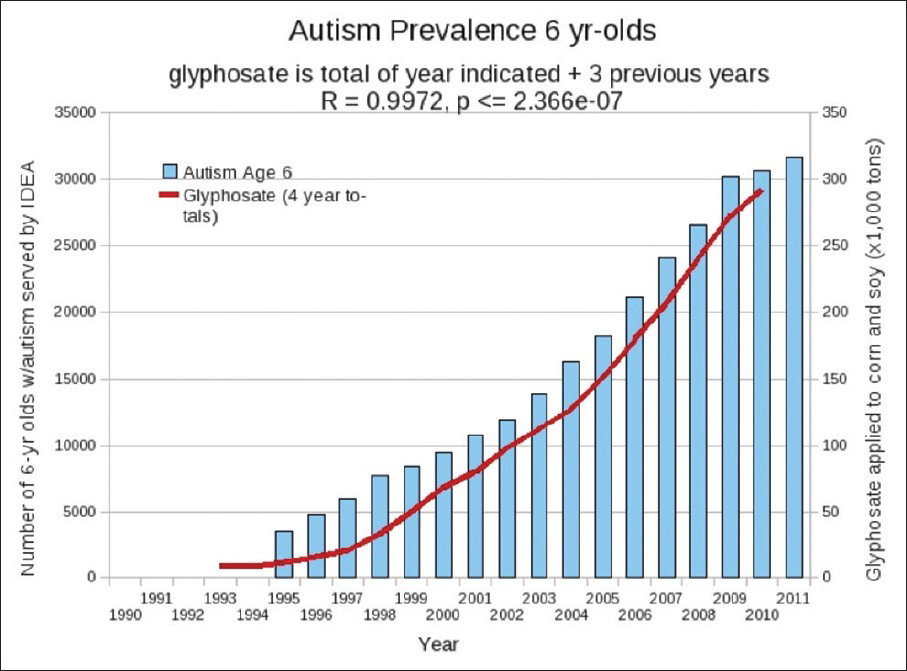
Figure 1
Plots of amount of glyphosate applied to corn and soy crops in the US over the previous 4 years (red), provided by the US Department of Agriculture, compared with number of children enrolled in the first grade in the public school system under the autism category according to the Individuals with Disabilities Education Act (IDEA) (blue bars). (Figure courtesy of Dr. Nancy Swanson)
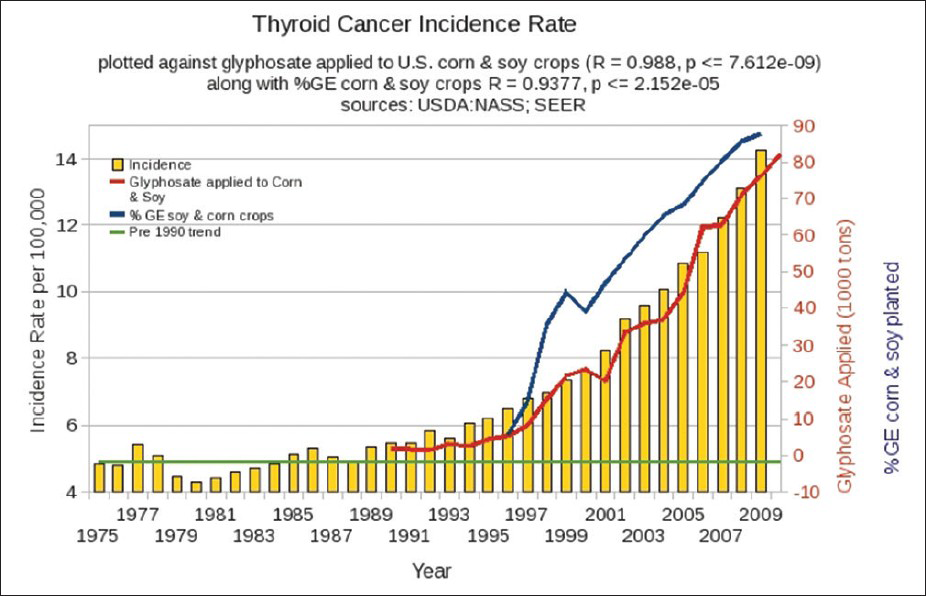
Thyroid dysfunction can be predicted as well, and low maternal thyroid function predicts autism in the fetus.[
Sprague-Dawley rats fed a Mn-deficient diet had significantly reduced concentrations of Mn in liver, kidney, heart, and pancreas, compared with controls.[
Much remains elusive about Mn's roles in cellular metabolism, but it is clear that it is very important. For instance, Target of Rapamycin Complex 1 (TORC1) accelerates the aging process in cells from yeast to mammals,[
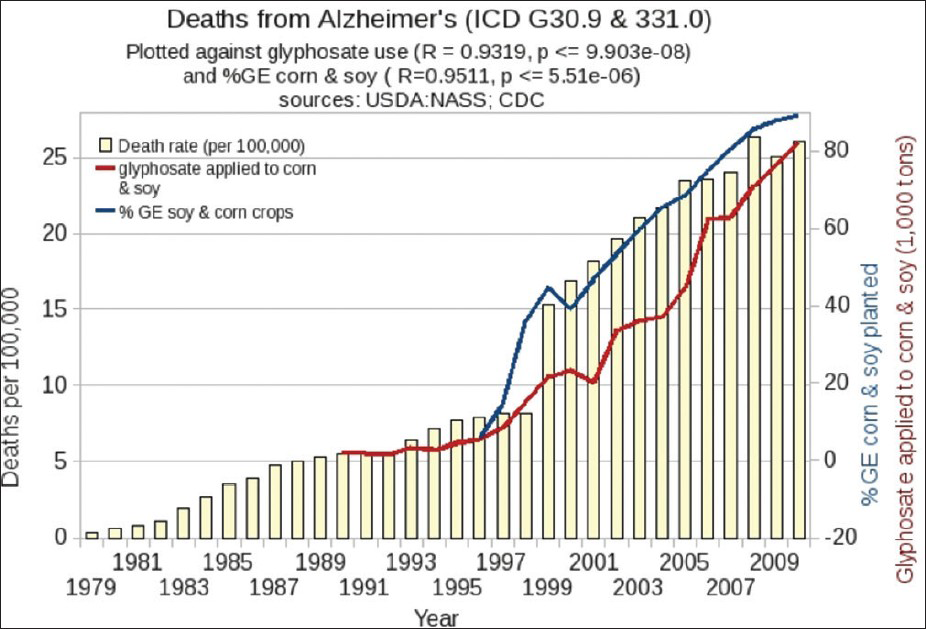
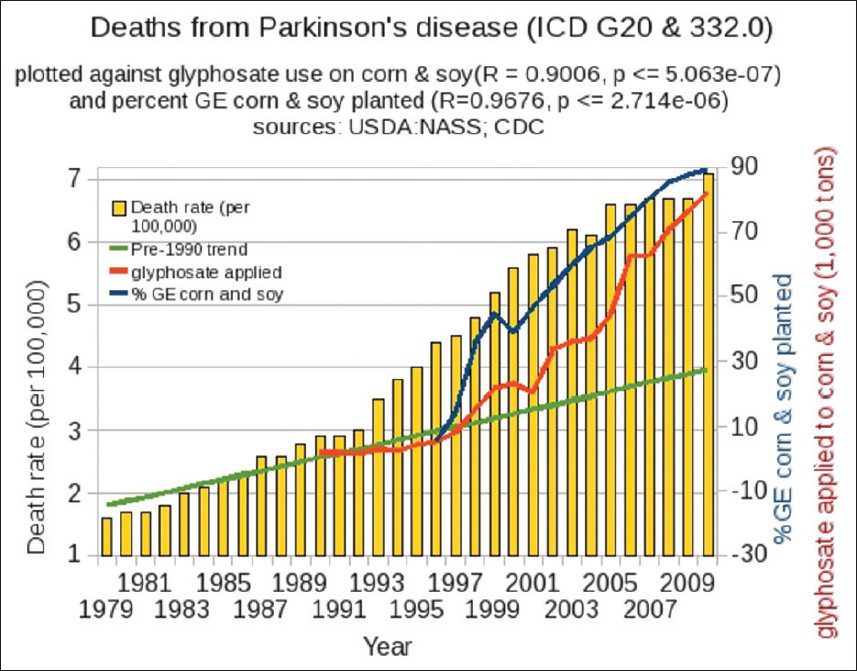
We infer, paradoxically, that both Mn deficiency and Mn toxicity, attributable to glyphosate, can occur simultaneously. Because of glyphosate's disruption of CYP enzymes, the liver becomes impaired in its ability to dispose of Mn via the bile acids, and instead it transports the Mn via the vagus nerve to brainstem nuclei, where excess Mn leads to PD. Recently, PD has also increased dramatically, in step with glyphosate usage on corn and soy [
Ironically, while the brainstem suffers from excess Mn, the rest of the brain incurs Mn deficiency due to the depressed serum levels of Mn. Mn is particularly important in the hippocampus, and deficiency there can lead to seizures. A high incidence of seizures is found in children with autism.[
Mn uptake in the brain is normally enhanced during the neonatal period in rats, and proper development of the hippocampus depends on Mn.[
Another mechanism by which glyphosate in soy formula could cause seizures is through bilirubin production. Serum concentrations of bilirubin were elevated in catfish exposed to sublethal doses of Roundup®, in a dose-dependent relationship.[
GLYPHOSATE AND MICROBIAL ANTIBIOTIC INTOLERANCE
Microbial antibiotic tolerance and resistance are a growing problem worldwide, likely fueled by horizontal gene transfer among different bacterial species.[
Studies have shown that increased mutation rates due to chronic low level exposure to one antibiotic can induce an accelerated rate of development of resistance to diverse other antibiotics.[
Glyphosate has also been demonstrated as a remarkable antimicrobial synergist. It greatly increases the cidal effects of other antimicrobials, particularly when combined as salts of glyphosate. A concentration dependent synergy index (SI) ranging from 0.34 to 5.13 has been recorded for the Zn salt of glyphosate.[
Glyphosate acts as a catalyst for the development of antibiotic resistance genes in pathogens. Since both poultry and cow manure are used as natural fertilizers in crops, it can be expected that a vector for microbial resistance to multiple drugs is through contamination of fruits and vegetables. Indeed, multiple resistance genes have been identified from diverse phyla found in cow manure, including Proteobacteria, Firmicutes, Bacteroidetes, and Actinobacteria, that is, in phylogenetically diverse organisms.[
One of the ways in which glyphosate is toxic to plants is through disruption of chlorophyll synthesis, due to suppression of the activity of the first enzyme in pyrrole synthesis.[
Thus, glyphosate can be expected to disrupt synthesis of these biologically essential molecules. Pseudomonas normally thrives in the small bowel and produces abundant cobalamin that may be a significant source for the human host.[
A key component of glyphosate's action is its ability to chelate minerals, particularly transition metals such as Mn. Glyphosate forms strong complexes with the transition metals via the amino, the carboxylic, and the phosphonic moieties in the molecule. Each of these can coordinate separately to metal ions or in combination as bidentate or tridentate ligands.[
ANALOGY WITH ARSENIC AND ALUMINUM
Chronic kidney disease is clearly associated with multiple environmental toxicants.[
Arsenic is normally disposed of by the liver through biliary excretion. In rats exposed to arsenic, large amounts of GSH appeared in the bile simultaneously with biliary excretion of arsenic.[
Glyphosate also chelates aluminum,[
However, it has been demonstrated through experimentation that glyphosate prefers divalent cations. Thus, aluminum would enter the bloodstream via the digestive tract's portal vein to the organs traveling with albumin, which is known to attach and transport many xenobiotics. It is well established that citrate also binds aluminum and promotes its uptake past the gut barrier through a mechanism that parallels glyphosate's binding to aluminum.[
Considering these observations regarding aluminum and arsenic, it is reasonable to expect that something similar might happen with Mn. Unlike these other two, however, Mn plays many essential roles in the body, and so its chelation by glyphosate would interfere with its bioavailability in the general circulation. Just as for arsenic, bile acids play a critical role in Mn homeostasis. Bile is the major excretory route of injected Mn.[
Glyphosate, a dipolar zwitterion, is toxic in part due to its bio-transformative properties as pH varies. We postulate that Mn, which is transported in the blood stream bound to glyphosate, is oxidized to Mn3+ following its release in the localized acidic environment of sulfated glycosaminoglycans (GAGs) in the glycocalyx lining the capillary wall.[
Mn is a transition metal, and therefore it can catalyze oxidative reactions in neurons via the Fenton reaction.[
It has been shown that glyphosate enhances the oxidation of Mn from a 2+ oxidation state to 3+, both in solution and on an inert surface.[
We suggest that another route for Mn transport is the vagus nerve, which delivers Mn from the liver to the brainstem nuclei. When bile acid synthesis is impaired, the brainstem nuclei can acquire neurotoxic levels of Mn, while serum levels are simultaneously depressed.
GUT BACTERIA DYSBIOSIS AND ANXIETY
Anxiety disorder is a comorbidity of autism,[
Glyphosate has been shown to severely deplete Mn uptake by plants, both by the roots and by the shoots.[
Certain species of gut bacteria, such as members of the Lactobacillus family, utilize Mn in novel ways for protection from oxidation damage, and, as a consequence, their requirements for Mn are much higher than those of other species.[
The liver regulates the amount of Mn in the general vascular circulation, by incorporating any excess into bile acids, which gives the gut bacteria repeated chances to take it up. However, production of bile acids depends upon CYP enzymes, which are disrupted by glyphosate.[
Lactobacillus tends to reside in the foregut.[
A recent study demonstrated that Roundup® in concentrations lower than those recommended in agriculture inhibited microbial growth of three microorganisms that are widely used as starters in fermentation of milk products,[
Bruno-Bárcena et al.[
Several members of the Lactobacillus family are capable of producing the inhibitory neurotransmitter γ-aminobutyric acid (GABA) via the enzyme glutamate decarboxylase, and this may be a reason for their ability to improve symptoms of anxiety. Experiments with Lactobacillus probiotics in mice demonstrated neurochemical and behavioral effects related to changes in GABAergic expression in regions of the brain that control mood.[
Patients suffering from chronic fatigue syndrome (CFS) often have imbalances in microbial flora,[
MN-SUPEROXIDE DISMUTASE AND MITOCHONDRIAL DYSFUNCTION
Mitochondrial dysfunction, particularly for the neutrophils, which perform important immune system functions, is implicated in CFS,[
Recent experiments on goldfish involved exposing them for 96 h to Roundup® at concentrations ranging from 2.5 to 20 mg/L.[
A recent study on the fish species Anguilla anguilla exposed to environmentally realistic levels of glyphosate over a short time period revealed DNA strand breaks in both liver and gills, along with a suppression of SOD activity in the liver.[
Aconitase, an enzyme that converts citrate to isocitrate, is a crucial participant in Complex I of the citric acid cycle in the mitochondria. Many neurodegenerative diseases have been linked to decreased aconitase activity due to oxidative stress, including Friedreich ataxia,[
A postmortem study of brains of autistic individuals showed a striking decrease in aconitase activity in the cerebellum associated with a similar decrease in glutathione redox antioxidant capacity (GSH/GSSG), with the plot producing a near 100% separation between cases and controls.[
In another study, cells from children with autism exhibited higher oxidative stress than control cells, including a 1.6-fold increase in reactive oxygen species (ROS) production, 1.5-fold increase in mitochondrial DNA copy number per cell, and more deletions.[
OXALATE
Monsanto's Roundup herbicide, WeatherMax® with Transorb II Technology, is now a preferred formulation and uses a potassium salt of glyphosate (48.8%). The Transorb Technology, which utilizes a dual surfactant system and adjuvants, was first introduced in 1996 as Roundup ULTRA. In 1998, the formula was altered and released as WeatherMAX, then further improved and released in 2005 as the current product.
The inert ingredients have undergone several changes over the years to include formula variations, which included the use of dual surfactants of siloxane copolymers and Polyoxyethylated tallow amine (POEA). As noted by Monsanto, “Promotion of stomatal infiltration of glyphosate by an organosilicone surfactant reduces the critical rainfall period,” hence the rain-fastness of Roundup WeatherMax® with Transorb® 2 Technology. In 1995, US Patent #5,464,806 was issued which reflected another product formulation improvement and move from the use of Silwet-77 siloxane surfactant due to phase separation problems, to the use of an acetylenic diol.[
This modification may be related to the recent increase in health issues concerning excess serum oxalate in the United States and elsewhere, linked to both autism and kidney stones. A study comparing children with autism with controls found a 3-fold increase in serum oxalate levels in the children with autism,[
Both the high oxalate content of the soy and the high serum oxalate in humans could be due to impaired oxalate metabolism. Oxalate metabolism by oxalate oxidase in plants and by oxalate decarboxylases in fungi and a few bacteria, such as Bacillus subtilis, are both dependent on Mn as a cofactor.[
The sulfate ion transporter, Sat-1, plays an important role not only in sulfate transport but also in oxalate transport,[
The conversion of glyoxylate to oxalate by the enzyme lactate dehydrogenase is inhibited by oxalate.[
Aside from the obvious damaging effects of oxalate crystals on tissues, the oxalate, whose metabolism is impaired due to Mn deficiency, will also interfere with the metabolism of glyphosate, likely greatly increasing both its effectiveness as an herbicide and its toxicity to mammals. Under oxalate stress conditions, both superoxide and the hydroxyl radical are produced in excess amounts.[
AMMONIA, GLUTAMATE, AND NEUROTOXICITY
In this section, we will show that both glutamate and ammonia are implicated as neurotoxins in connection with autism and other neurological diseases, and we will offer the simple explanation that Mn deficiency leads to impaired activity of glutamine synthase and arginase, both of which utilize Mn as a cofactor. Mn deficiency can also explain the increased risk to epilepsy found in autism, due to the fact that Mn decreases T2 relaxation time. Mn-deprived rats are more susceptible to convulsions.[
Blaylock and Strunecká[
The neurotransmitter glutamate has been implicated as an excitatory neurotoxin in the brain not only in autism but also in association with multiple neurological diseases, including AD, PD, amyotrophic lateral sclerosis (ALS), and multiple sclerosis.[
TNF-α, secreted by activated microglia in the brain, is a major cytokine leading to neurotoxicity in association with multiple neurological diseases. A major component of the damaging effect of TNF-α is the autocrine induction of the release of glutamate from microglia.[
Glutamine synthase depends upon Mn as a cofactor, so depleted Mn supplies would lead to a build-up of glutamate that cannot be returned to the neurons using normal channels. Multiple sclerosis is associated with both depleted Mn in the cerebrospinal fluid[
Further confirmation of glutamate dysbiosis in autism comes from a study on levels of 25 amino acids in the platelet-poor plasma of high-functioning autistic children compared with normal controls, which revealed that only glutamate and glutamine were abnormally expressed in the children with autism, with a highly significant (P < 0.002) excess of glutamate and a highly significant (P < 0.004) decrease in glutamine.[
It is intriguing to us that Mn deficiency leads to a pair of complementary pathologies – excess glutamate along with aconitase deficiency, which together allow for the cells to generate ATP by metabolizing glutamate instead of glucose. Glutamate enters the citric acid cycle beyond Complex I, thus bypassing the step that is impaired by aconitase deficiency. This is a rather elegant regulatory system that provides energy even in the face of Complex I impairment.
The prevalence of epilepsy in the US is similar to that of diabetes, making it a common disorder affecting 2 million Americans.[
Ammonia is a well-established neurotoxin, which accumulates when the urea cycle is unable to keep up with ammonia released from protein breakdown.[
Excess ammonia due to impaired ability to detoxify excess nitrogen via the urea cycle can lead to impaired memory, shortened attention span, sleep–wake cycle disruption, brain edema, intracranial hypertension, seizures, ataxia, and coma.[
A case study of a 4-year-old girl showed that a genetic defect in arginase could present as pervasive developmental delay not otherwise specified (PDD-NOS) with many similarities to autism.[
Seneff et al.[
CHOLESTASIS AND BILIRUBINEMIA
Mn stimulates cholesterol synthesis in the liver, presumably because bile acids, derived from cholesterol, are needed to bind to Mn and carry it back to the gut for recycling.[
Studies with rats have shown that Lactobacillus plantarum probiotic supplements lower serum LDL levels.[
Cholestasis is a blockage of bile acid flow, which often occurs as a side effect of various pharmaceutical drugs.[
Inflammatory bowel disease (colitis and Crohn's disease) has been increasing in frequency in the US over the past 20 years, in step with glyphosate usage on corn and soy crops, as shown in [
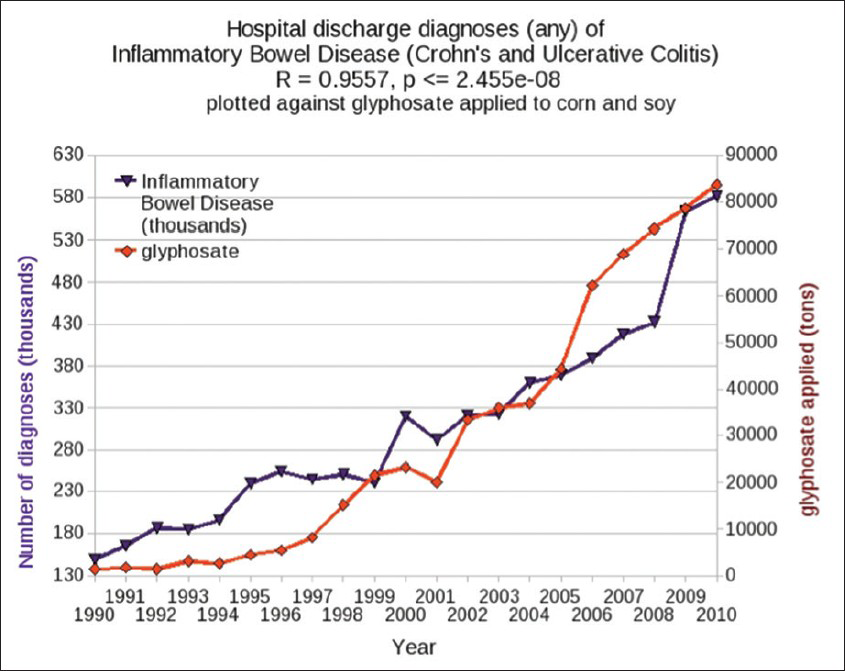
Ironically, while Mn-SOD depends upon Mn as a cofactor, excess exposure to Mn can inhibit SOD expression. Experiments exposing rats to excess Mn revealed several pathological effects on the liver, including inhibition of SOD and GSH peroxidase, as well as decreased levels of GSH and reduced levels of sodium-potassium ATPase activity.[
Bilirubinemia in neonates is a risk factor for autism, particularly when it is unbound and unconjugated.[
G6PD is the main enzyme responsible for regeneration of NADPH, an essential requirement for GSH reductase activity.[
A mouse model of cholestasis can be induced by exposing mice to Mn, followed shortly by exposure to bilirubin.[
The incorporation of cholesterol products into exported bile acids is crucial for regulating cholesterol homeostasis.[
Intracellular accumulation of bile acids, associated with cholestasis, is known to be toxic to hepatocytes. The accumulation of bile acids in the cholestatic liver induces oxidative stress and apoptosis, resulting in damage to the liver parenchyma, and, ultimately, extrahepatic tissues as well.[
On the other hand, the hydrophilic bile acid, ursodeoxycholic acid (UDCA), is protective, and its protective effects have been proposed to be due to its ability to induce the expression of CYP3A4, a bile-metabolizing enzyme, in hepatocytes.[
Another enzyme class that was discovered to be downregulated in the liver by glyphosate in the goldfish study is GST, an important class of enzymes. The main function of the GST enzymes is the detoxification of electrophilic xenobiotics by GSH conjugation.[
One final factor in cholestasis is vitamin K deficiency, which is associated with cholestatic liver disease.[
SALMONELLA INFECTION
Multiple drug-resistant strains of Salmonella are becoming an increasing problem both in the industrialized[
Our research into the pathology of Salmonella has uncovered a complex interplay of many factors that may be responsible for the epidemic, which includes an important role for Mn, as well as a role for industrial processing of lecithin from soy, and bile acid disruption by glyphosate. Commercial lecithin is a mixture of phospholipids and various metabolites, often derived from soy. In food processing, phospholipase A (PLA) is applied enzymatically to hydrolyze phospholipids in lecithin into lysophospholipids and fatty acids, in order to improve its emulsification, surfactant, and lubricant properties.[
Salmonella depend on cyclic adenosine monophosphate (cAMP) for flagella formation, and therefore for motility.[
Salmonella are remarkably resilient in an inflammatory environment, and, in a novel strategy for survival in a hostile environment, they take advantage of tetrathionate produced from oxidation of thiosulfate by ROS as a terminal electron acceptor in processing ethanolamine derived from host lysophospholipids as a source of energy not available to other microbes.[
It has only recently been appreciated that Mn plays an important role in the virulence of Salmonella.[
A feature unique to Salmonella is that they are especially adept at binding to cholesterol in the gall bladder, particularly in association with gallstones, and they also possess adaptive responses that allow them to survive the harsh environment of the bile acid milieu, as well as the highly acidic environment of the stomach.[
Several different species of Salmonella can form biofilms on human gallstones, which is dependent upon the presence of bile.[
Fibrates are hypolipidemic agents that are known to suppress bile acid synthesis via suppression of peroxisome proliferator-activated receptor γ (PPAR-γ).[
CHONDROITIN SULFATE, OSTEOMALACIA AND ARTHRITIS
Vitamin D deficiency has reached epidemic proportions in the US and increasingly around the world in recent years.[
Samsel and Seneff[
Osteoarthritis is another pathology likely related to Mn deficiency, impaired chondroitin sulfate synthesis and impaired vitamin D activation. Vitamin D deficiency is associated with rheumatoid arthritis.[
A mysterious epidemic of a new disease, called “sea star wasting syndrome,” is currently sweeping across the Pacific coast of North America, affecting at least 12 different species of sea stars.[
A protective layer of mucopolysaccharides called mucus is secreted by corals, and it has been characterized as containing sulfated glycoproteins similar to chondroitin sulfate,[
PARKINSON'S DISEASE
It might be anticipated that a simple Mn mineral supplement could correct for the problem of glyphosate's chelation of Mn, but this assumption is almost certainly false. We suggest that glyphosate's disruption of Mn homeostasis leads to extreme sensitivity to Mn bioavailability, making it easy to err in the direction of either too little or too much.[
The fact that death from PD has increased in the past two decades in step with glyphosate usage on corn and soy suggests that there may be a role for glyphosate to play in Parkinson's pathology [
An experiment where rats were exposed to Mn via intranasal instillation demonstrated that the Mn could enter the brain through the olfactory bulbs by following major neuronal pathways.[
Elder et al.[
Manganism is distinct from PD mainly in that it is the locus coeruleus that is preferentially damaged rather than the substantia nigra.[
Berthoud et al.[
A study using MRI technology to detect Mn distribution in the brain in marmosets and rats following Mn injections revealed an accumulation in the basal ganglia as well as other parts of the brain that were situated near the ventricles, suggesting redistribution via the cerebrospinal fluid.[
Both the substantia nigra and the locus coeruleus are characterized by high concentrations of neuromelanin. While it is unclear what role neuromelanin plays, one hypothesis is that it carries a protective action through its unique ability to accumulate and retain various amines and metallic cations, especially Mn.[
Glyphosate itself likely also contributes directly to PD. PD is caused by degeneration of dopaminergic neurons in the substantia nigra, attributed to mitochondrial dysfunction, oxidative stress, and protein aggregation.[
A study on serum Mn levels in infants and their association with neurodevelopment revealed a U-shaped curve, with both too little and too much Mn leading to impaired development.[
Dopamine suppresses thyroid stimulating hormone, and therefore dopamine insufficiency can lead to overactive thyroid and potential burnout of the thyroid gland.[
PRION DISEASES AND MANGANESE
Prions are a special class of proteins that acquire alternative conformations that can become self-propagating. There is a class of neurological diseases called transmissible spongiform encephalopathies that are believed to be caused by misfolding of prion proteins, predominantly accumulating in the gray matter in the brain.[
Prion proteins have been shown to bind Cu in vivo,[
The possible link between excess Mn and prion diseases was also supported in a later study by Masánová et al.,[
Mn-SOD-/- mice die between days 3 and 13 following birth. They exhibit a marked dilated cardiomyopathy, neurodegeneration and fatty liver disease.[
Purdey[
New experimental data support the idea that the class of prion diseases should be expanded to include AD, PD, and related tauopathies.[
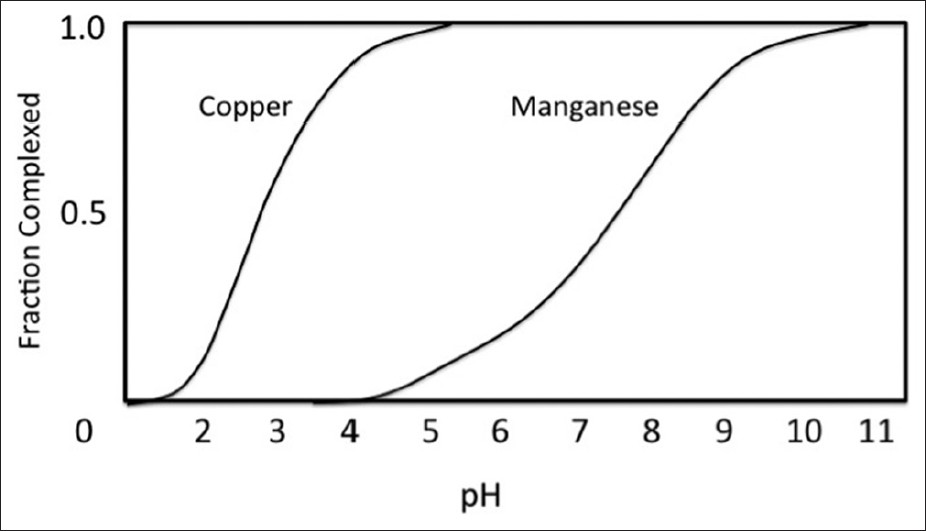
Figure 8
Fractions of metal complexed with glyphosate as a function of pH, for copper and manganese. Figure adapted from Lundager Madsen et al[
MN AND INFERTILITY
Multiple papers on rodent studies have indicated disruption of the male reproductive system by glyphosate. Acute treatment of 60-day-old male Sprague-Dawley rats to Roundup® caused a marked increase in aromatase mRNA in testicular tissue,[
In vitro studies on Sertoli cells from mouse testis demonstrated that glyphosate opens L-type voltage-dependent calcium channels, leading to calcium-overload cell death.[
EVIDENCE OF EXPOSURE
Glyphosate-containing herbicides are applied to crops several times each season both for killing weeds and for desiccation just prior to the harvest in non-Roundup®-Ready crops, such as wheat and sugar cane. It accumulates in the leaves, grains, and fruit, and thus cannot be removed by washing and, furthermore, is not broken down by cooking.[
A recently published study by Shehata et al.[
A recent paper by Nevison[
There has been very little testing of glyphosate levels in either humans or other mammals. Glyphosate is passed in both the urine and the feces. Recent research from Europe has shown that glyphosate is consistently present in significant amounts in the urine of cows consuming Roundup®-Ready feed, as well as in the organs and meat of cattle.[
CONCLUSION
Glyphosate is the most widely used herbicide on the planet, in part because of its perceived low toxicity to humans. In this paper, we propose that glyphosate's chelation of Mn, working together with other known effects of glyphosate such as CYP enzyme suppression and depletion of derivatives of the shikimate pathway in microorganisms, may explain the recent increase in incidence of multiple neurological diseases and other pathologies. We have shown that glyphosate's disruption of Mn homeostasis can lead to extreme sensitivity to variations in Mn bioavailability: While Mn deficiency in the blood leads to impairment of several Mn dependent enzymes, in contrast, excess Mn readily accumulates in the liver and in the brainstem due to the liver's impaired ability to export it in the bile acids. This pathology can lead to liver damage and PD. Mn depletion in the gut due to chelation by glyphosate selectively affects Lactobacillus, leading to increased anxiety via the gut–brain access. Both low Lactobacillus levels in the gut and anxiety syndrome are known features of autism, and Lactobacillus probiotic treatments have been shown to alleviate anxiety. Increased incidence of Salmonella poisoning can also be attributed to glyphosate, through its impairment of bile acid synthesis. Low Mn bioavailability from the blood supply to the brain leads to impaired function of glutamine synthase and a build-up of glutamate and ammonia in the brain, both of which are neurotoxic. Excess brain glutamate and ammonia are associated with many neurological diseases. At the same time, impaired function of Mn-SOD in the mitochondria results in mitochondrial damage, also a hallmark of many neurological diseases. Mn deficiency can account for poor sperm motility and therefore low fertilization rates, as well as poor bone development leading to osteoporosis and osteomalacia. Sea star wasting syndrome and the collapse of coral reefs may in fact be an ecological consequence of the environmental pervasiveness of the herbicide. Many diseases and conditions are currently on the rise in step with glyphosate usage in agriculture, particularly on GM crops of corn and soy. These include autism, AD, PD, anxiety disorder, osteoporosis, inflammatory bowel disease, renal lithiasis, osteomalacia, cholestasis, thyroid dysfunction, and infertility. All of these conditions can be substantially explained by the dysregulation of Mn utilization in the body due to glyphosate.
ACKNOWLEDGMENTS
This work was funded in part by Quanta Computers, Taipei, Taiwan, under the auspices of the Qmulus Project. The authors thank Dr. Nancy Swanson for providing the plots showing correlations over time of multiple diseases and conditions with glyphosate usage on corn and soy in the US.
Commentary
- Gene Expression and Therapy Group, King's College London, Faculty of Life Sciences and Medicine, Department of Medical and Molecular Genetics, 8th Floor, Tower Wing, Guy's Hospital, Great Maze Pond, London, SE1 9RT, UK. E-mail: *Michael N. Antoniou -
michael.antoniou@kcl.ac.uk
Glyphosate-based herbicides (GBH) are the major pesticides used worldwide. Initially patented as a metal ion chelator, glyphosate rapidly jumped to a leading position as an active ingredient of commercial pesticides from the 1970s when Monsanto discovered its herbicidal activities. The herbicidal mode of action of glyphosate is primarily to inhibit the shikimic acid pathway,[
Glyphosate is an aminophosphonic analog of glycine. The fact that glycine and other amino acids like glutamate function as neurotransmitters and play a crucial role in brain function, makes the potential neurotoxic effects of glyphosate a matter of concern.[
The most recent reevaluation of the acceptable daily intake (ADI) for glyphosate within the European Union (EU) conducted by the German regulatory agency (BfR), states that this has been determined by scrutiny of approximately 450 regulatory toxicological studies and 900 publications from the scientific literature.[
However, a debate continues as to the soundness of this BfR-led assessment as the studies taken into account were performed with an experimental design adapted to the study of poisoning effects based on the principle of “the dose makes the poison”; that is, the higher the dose the greater the poisoning effect. Of major concern is that none of the studies referred to incorporated testing principles derived from a contemporary understanding of (neuro) endocrinology or developmental epigenetics.
In the classic theory of toxicology, as applied for the study of glyphosate toxicity at a regulatory level, a nontoxic threshold is evidenced by decreasing the level of exposure and assuming that toxic effects observed are a linear response to the dose. Lower doses corresponding to environmental exposures are assumed to be safe and are not tested. However, in contrast to a classical poison, an endocrine disruptive chemical (EDC) will alter the functioning of hormonal systems and induce adverse effects at various dosage levels. Such EDC effects will, in some cases, occur in a nonlinear (non-monotonic) manner at environmentally relevant levels of exposure and will not be observed at higher doses. In addition and not surprisingly, EDC effects can be sex-specific in nature. Such nonmonotonic and sex-specific EDC effects have been extensively described for common pollutants.[
Major endpoints of toxicity such as neurodevelopmental, reproductive, and transgenerational effects in humans still needs to be investigated at the glyphosate ADI and other concentrations reflective of human levels of exposure. Given its increasing wide-scale use and consequent rise in exposure, we urgently call for greater research efforts on the toxicology of glyphosate and its commercial herbicide formulations, as well as pesticide neurodevelopmental effects in general. Furthermore, pesticide combinatorial (additive or synergistic) effects remain a poorly investigated subject and area of concern that needs to be addressed. Such studies are particularly relevant to the brain since it is physiologically dependent on neurosteroids, making it potentially very sensitive to endocrine disruptive compounds.
References
1. Abdel-Megeed A, Sadik MW, Al-Shahrani HO, Ali HM. Phyto-microbial degradation of glyphosate in Riyadh area. Int J Microbiol Res. 2013. 5: 458-
2. Adams JB, Audhya T, McDonough-Means S, Rubin RA, Quig D, Geis E. Nutritional and metabolic status of children with autism vs. neurotypical children, and the association with autism severity. Nutr Metab. 2011. 8: 34-
3. Afzal-Ahmed I, Mann GE, Shennan AH, Poston L, Naftalin RJ. Preeclampsia inactivates glucose-6-phosphate dehydrogenase and impairs the redox status of erythrocytes and fetal endothelial cells. Free Radic Biol Med. 2007. 42: 1781-90
4. Aguirre JD, Clark HM, McIlvin M, Vazquez C, Palmere SL, Grab DJ. A manganese-rich environment supports superoxide dismutase activity in a Lyme disease pathogen, Borrelia burgdorferi. J Biol Chem. 2013. 288: 8468-78
5. Akoume MY, Perwaiz S, Yousef IM, Plaa GL. Synergistic role of 3-hydroxy-3-methylglutaryl coenzyme A reductase and cholesterol 7α-hydroxylase in the pathogenesis of manganese-bilirubininduced cholestasis in rats. Toxicol Sci. 2003. 73: 378-85
6. Albert MJ, Mathan VI, Baker SJ. Vitamin B12 synthesis by human small intestinal bacteria. Nature. 1980. 283: 781-2
7. Allocati N, Federici L, Masulli M, Di Ilio C. Glutathione transferases in bacteria. FEBS J. 2009. 276: 58-75
8. Ali MM, Newsom DL, González JF, Sabag-Daigle A, Stahl C, Steidley B. Fructose-asparagine is a primary nutrient during growth of Salmonella in the inflamed intestine. PLOS Pathogens. 2014. 10: e1004209-
9. Álvarez-Ordóñez A, Begley M, Prieto M, Messens W, López M, Bernardo A. Salmonella spp. survival strategies within the host gastrointestinal tract. Microbiology. 2011. 157: 3268-81
10. Amin SB, Smith T, Wang H. Is neonatal jaundice associated with autism spectrum disorders: A systematic review. J Autism Dev Disord. 2011. 41: 1455-63
11. Ananthanarayanan M, Balasubramanian N, Makishima M, Mangelsdorf DJ, Suchy FJ. Human bile salt export pump promoter is transactivated by the farnesoid X receptor/bile acid receptor. J Biol Chem. 2001. 276: 28857-65
12. Antoniou M, Habib ME, Howard CV, Jennings RC, Leifert C, Nodari RO. Teratogenic effects of glyphosate-based herbicides: Divergence of regulatory decisions from scientific evidence. J Environ Anal Toxicol. 2012. S4: 006-
13. Archibald FS, Duong MN. Manganese acquisition by Lactobacillus plantarum. J Bacteriol. 1984. 158: 1-8
14. Archibald FS, Fridovich I. Manganese, superoxide dismutase, and oxygen tolerance in some lactic acid bacteria. J Bacteriol. 1981. 146: 928-36
15. Attili AF, Angelico M, Cantafora A, Alvaro D, Capocaccia L. Bile acid- induced liver toxicity: Relation to the hydrophobic-hydrophilic balance of bile acids. Med Hypotheses. 1986. 19: 57-69
16. Avery D, Winokur G. The efficacy of electroconvulsive therapy and antidepressants in depression. Biol Psychiatry. 1977. 12: 507-23
17. Ballatori N, Miles E, Clarkson TW. Homeostatic control of manganese excretion in the neonatal rat. Am J Physiol. 1987. 252: R842-7
18. Baly DL, Curry DL, Keen CL, Hurley LS. Effect of manganese deficiency on insulin secretion and carbohydrate homeostasis in rats. J Nutr. 1984. 114: 1438-46
19. Baly DL, Schneiderman JS, Garcia-Welsh AL. Effect of manganese deficiency on insulin binding, glucose transport and metabolism in rat adipocytes. J Nutr. 1990. 120: 1075-9
20. Barbosa ER, Leiros da Costa MD, Bacheschi LA, Scaff M, Leite CC. Parkinsonism after glycine-derivate exposure. Mov Disord. 2001. 16: 565-8
21. Barrett KA, McBride MB. Oxidative degradation of glyphosate and aminomethyl-phosphonate by manganese oxide. Environ Sci Technol. 2005. 39: 9223-8
22. Battaglin WA, Meyer MT, Kuivila KM, Dietze JE. Glyphosate and its degradation product AMPA occur frequently and widely in U.S. soils, surface water, groundwater, and precipitation. J Am Water Resour Assoc. 2014. 50: 275-90
23. Bayliss EA, Hambidge KM, Sokol RJ, Stewart B, Lilly JR. Hepatic concentrations of zinc, copper and manganese in infants with extrahepatic biliary atresia. J Trace Elem Med Biol. 1995. 9: 40-3
24. Becker KG, Schultz ST. Similarities in features of autism and asthma and a possible link to acetaminophen use. Med Hypotheses. 2010. 74: 7-11
25. Becker KG. Autism, asthma, inflammation, and the hygiene hypothesis. Medl Hypotheses. 2007. 69: 731-40
26. Benbrook CM. Impacts of genetically engineered crops on pesticide use in the U.S. – the first sixteen years. Environ Sci Europe. 2012. 24: 24-
27. Bentley R, Meganathan R. Biosynthesis of Vitamin K (Menaquinone) in Bacteria. Microbiol Rev. 1982. 46: 241-80
28. Berk PD, Javitt NB. Hyperbilirubinemia and cholestasis. Am J Med. 1978. 64: 311-26
29. Bernards ML, Thelen KD, Penner D, Muthukumaran RB, McCracken JL. Glyphosate interaction with manganese in tank mixtures and its effect on glyphosate absorption and translocation. Weed Sci. 2005. 53: 787-94
30. Berthoud HR, Kressel M, Neuhuber WL. An anterograde tracing study of the vagal innervation of rat liver, portal vein and biliary system. Anat Embryol. 1992. 186: 431-42
31. Bertolotti M, Concari M, Loria P, Abate N, Pinetti A, Guicciardi ME. Effects of different phenotypes of hyperlipoproteinemia and of treatment with fibric acid derivative on the rates of cholesterol 7alpha-hydroxylation in humans. Arterioscler Thromb Vasc Biol. 1995. 15: 1064-9
32. Blaylock RL, Strunecka A. Immune-glutamatergic dysfunction as a central mechanism of the autism spectrum disorders. Curr Med Chem. 2009. 16: 157-70
33. Bock NA, Paiva FF, Nascimento GC, Newman JD, Silva AC. Cerebrospinal fluid to brain transport of manganese in a non-human primate revealed by MRI. Brain Res. 2008. 1198: 160-70
34. Bodnar LM, Klebanoff MA, Gernand AD, Platt RW, Parks WT, Catov JM. Maternal Vitamin D status and spontaneous preterm birth by placental histology in the US Collaborative Perinatal Project. Am J Epidemiol. 2014. 179: 168-76
35. Bøhn T, Cuhra M, Traavik T, Sanden M, Fagan J, Primicerio R. Compositional differences in soybeans on the market: Glyphosate accumulates in Roundup Ready GM soybeans. Food Chem. 2014. 153: 207-15
36. Bolze MS, Reeves RD, Lindbeck FE, Kemp SF, Elders MJ. Influence of manganese on growth, somatomedin and glycosaminoglycan metabolism. J Nutr. 1985. 115: 352-8
37. Bosoi CR, Rose CF. Identifying the direct effects of ammonia on the brain. Metab Brain Dis. 2009. 24: 95-102
38. Braak H, Del Tredici K, Rb U, De Vos RA, Ernst NH, Steur J. Staging of brain pathology related to sporadic Parkinsons disease. Neurobiol Aging. 2003. 24: 197-211
39. Braun T, Dods RF. Development of a Mn 2+ -sensitive, “soluble” adenylate cyclase in rat testis. Proc Nat Acad Sci USA. 1975. 72: 1097-101
40. Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, Dinan TG. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci U S A. 2011. 108: 16050-5
41. Brewster DW, Warren J, Hopkins WE. Metabolism of glyphosate in SpragueDawley rats: Tissue distribution, identification, and quantitation of glyphosate-derived materials following a single oral dose. Fundam Appl Toxicol. 1991. 17: 43-51
42. Briellmann RS, Syngeniotis A, Fleming S, Kalnins RM, Abbott DF, Jackson GD. Increased anterior temporal lobe T2 times in cases of hippocampal sclerosis: A multi-echo T2 relaxometry study at 3 T. AJNR Am J Neuroradiol. 2004. 25: 389-94
43. Brock AA, Chapman SA, Ulman EA, Wu G. Dietary manganese deficiency decreases rat hepatic arginase activity. J Nutr. 1994. 124: 340-4
44. Brown BE, Bythell JC. Perspectives on mucus secretion in reef corals. Mar Ecol Prog Ser. 2005. 296: 291-309
45. Brown DR, Hafiz F, Glasssmith LL, Wong BS, Jones IM, Clive C. Consequences of manganese replacement of copper for prion protein function and proteinase resistance. EMBO J. 2000. 19: 1180-6
46. Brown DR, Qin K, Herms JW, Madlung A, Manson J, Strome R. The cellular prion protein binds copper in vivo. Nature. 1997. 390: 684-7
47. Browne T, Holmes G. Epilepsy. N Engl J Med. 2001. 344: 1145-51
48. Bruno-Bárcena JM, Andrus JM, Libby SL, Klaenhammer TR, Hassan HM. Expression of a heterologous manganese superoxide dismutase gene in intestinal lactobacilli provides protection against hydrogen peroxide toxicity. Appl Environ Microbiol. 2004. 70: 4702-10
49. Cagnon L, Braissant O. Hyperammonemia-induced toxicity for the developing central nervous system. Brain Res Rev. 2007. 56: 183-97
50. Cakmak I, Yazici A, Tutus Y, Ozturk L. Glyphosate reduced seed and leaf concentrations of calcium, manganese, magnesium, and iron in non-glyphosate resistant soybean. Eur J Agron. 2009. 31: 114-9
51. Campbell IM, Robins DJ, Kelsey M, Bentley R. Biosynthesis of Bacterial Menaquinones (Vitamins K)$_2$). Biochemistry. 1971. 10: 3069-78
52. Cantu D, Fulton RE, Drechsel DA, Patel M. Mitochondrial aconitase knockdown attenuates paraquat-induced dopaminergic cell death via decreased cellular metabolism and release of iron and H 2 O 2. J Neurochem. 2011. 118: 79-92
53. Cappello M, Randazzo C, Bravatà I, Licata A, Peralta S, Craxì A. Liver function test abnormalities in patients with inflammatory bowel diseases: A hospital-based survey. Clin Med Insights Gastroenterol. 2014. 7: 25-31
54. Carl GF, Keen CL, Gallagher BB, Clegg MS, Littleton WH, Flannery DB. Association of low blood manganese concentrations with epilepsy. Neurology. 1986. 36: 1584-7
55. Carroll IM, Andrus JM, Bruno-Brcena JM, Klaenhammer TR, Hassan HM, Threadgill DS. Anti-inflammatory properties of Lactobacillus gasseri expressing manganese superoxide dismutase using the interleukin 10-deficient mouse model of colitis. Physiol Gastrointest Liver Physiol. 2007. 293: G729-38
56. Cassault-Meyer E, Gress S, Séralini GE, Galeraud-Denis I. An acute exposure to glyphosate-based herbicide alters aromatase levels in testis and sperm nuclear quality. Environ Toxicol Pharmacol. 2014. 38: 131-40
57. Castellani R, Smith MA, Richey GL, Perry G. Glycoxidation and oxidative stress in Parkinson disease and diffuse Lewy body disease. Brain Res. 1996. 737: 195-200
58. Castle LA, Siehl DL, Gorton R, Patten PA, Chen YH, Bertain S. Discovery and directed evolution of a glyphosate tolerance gene. Science. 2004. 304: 1151-4
59. Cattani D, de Liz Oliveira Cavalli VL, Heinz Rieg CE, Domingues JT, Dal-Cim T, Tasca CI. Mechanisms underlying the neurotoxicity induced by glyphosate-based herbicide in immature rat hippocampus: Involvement of glutamate excitotoxicity. Toxicology. 2014. 320C: 34-45
60. Cavuşoğlu K, Yapar K, Oruç E, Yalçın E. Protective effect of Ginkgo biloba L. leaf extract against glyphosate toxicity in Swiss albino mice. J Med Food. 2011. 14: 1263-72
61. Cebeci O, Budak H. Global expression patterns of three Festuca species exposed to different doses of glyphosate using the affymetrix genechip wheat genome array. Comp Funct Genomics 2009. 2009. p.
62. Centeno JA, Mullick FG, Martinez L, Page NP, Gibb H, Longfellow D. Pathology related to chronic arsenic exposure. Environ Health Perspect. 2002. 110: S883-6
63. Cerdeira AL, Duke SO. The current status and environmental impacts of glyphosate-resistant crops: A review. J Environ Qual. 2006. 35: 1633-58
64. Chan P, Mahler J. NTP technical report on the toxicity studies of glyphosate (CAS No. 1071-83-6) administered in dosed feed to F344/N rats and B6C3F1 mice. Toxic Rep Ser. 1992. 16: 1-D3
65. Chen J, Liu J, Zhang Y, Ye Z, Wang S. Decreased renal vitamin K-dependent gamma-glutamyl carboxylase activity in calcium oxalate calculi patients. Chin Med J (Engl). 2003. 116: 569-72
66. Clair E, Linn L, Travert C, Amiel C, Sralini GE, Panoff JM. Effects of Roundup() and glyphosate on three food microorganisms: Geotrichum candidum, Lactococcus lactis subsp. cremoris and Lactobacillus delbrueckii subsp. bulgaricus. Curr Microbiol. 2012. 64: 486-91
67. Claus Henn B, Ettinger AS, Schwartz J, Téllez-Rojo MM, Lamadrid-Figueroa H, Hernández-Avila M. Early postnatal blood manganese levels and childrens’ neurodevelopment. Epidemiology. 2010. 21: 433-9
68. Coburn JW, Mischel MG, Goodman WG, Salusky IB. Calcium citrate markedly enhances aluminum absorption from aluminum hydroxide. Am J Kidney Dis. 1991. 17: 708-11
69. Cole DJ, Grossbard E, Atkinson D.editors. Mode of action of glyphosate - A literature analysis. The herbicide glyphosate. London: Butterworths; 1985. p. 48-74
70. Collins SM, Surette M, Bercik P. The interplay between the intestinal microbiota and the brain. Nat Rev Microbiol. 2012. 10: 735-42
71. Cooper AJ. 13N as a tracer for studying glutamate metabolism. Neurochem Int. 2011. 59: 456-64
72. Cotzias GC, Papavasiliou PS, Hughes ER, Tang L, Borg DC. Slow turnover of manganese in active rheumatoid arthritis accelerated by prednisone. J Clin Invest. 1968. 47: 992-1001
73. Cox LM, Yamanishi S, Sohn J, Alekseyenko AV, Leung JM, Cho I. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell. 2014. 158: 705-21
74. Crawford RW, Reeve KE, Gunn JS. Flagellated but not hyperfimbriated Salmonella enterica serovar Typhimurium attaches to and forms biofilms on cholesterol-coated surfaces. J Bacteriol. 2010. 192: 2981-90
75. Cryan JF, Dinan TG. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. 2012. 13: 701-12
76. Curtis D, Sparrow R, Brennan L, Van der Weyden MB. Elevated serum homocysteine as a predictor for vitamin B12 or folate deficiency. Eur J Haematol. 1994. 52: 227-32
77. Daniell WE, Stockbridge HL, Labbe RF, Woods JS, Anderson KE, Bissell DM. Environmental chemical exposures and disturbances of heme synthesis. Environ Health Perspect. 1997. 105: 37-53
78. Daoust A, Barbier EL, Bohic S. Manganese enhanced MRI in rat hippocampus: A correlative study with synchrotron X-ray microprobe. Neuroimage. 2013. 64: 10-8
79. Dauer W, Przedborski S. Parkinsons disease: Mechanisms and models. Neuron. 2003. 39: 889-909
80. de Carmo Langiano V, Martinez CB. Toxicity and effects of a glyphosate-based herbicide on the Neotropical fish Prochilodus lineatus. Comp Biochem Physiol Part C. 2008. 147: 222-31
81. De Keersmaecker SC, Verhoeven TL, Desair J, Marchal K, Vanderleyden J, Nagy I. Strong antimicrobial activity of Lactobacillus rhamnosus GG against Salmonella typhimurium is due to accumulation of lactic acid. FEMS Microbiol Lett. 2006. 259: 89-96
82. de Lamirande E, Tuchweber B, Plaa GL. Morphological aspects of manganese-bilirubin induced cholestasis. Liver. 1982. 2: 22-7
83. de Liz Oliveira Cavalli LV, Cattani D, Heinz Rieg CE, Pierozan P, Zanatta L, Benedetti Parisotto E. Roundup disrupts male reproductive functions by triggering calcium-mediated cell death in rat testis and Sertoli cells. Free Radic Biol Med. 2013. 65: 335-46
84. DeLong GR. Autism, amnesia, hippocampus, and learning. Neurosci Biobehav Rev. 1992. 16: 63-70
85. De Roos AJ, Zahm SH, Cantor KP, Weisenburger DD, Holmes FF, Burmeister L. Integrative assessment of multiple pesticides as risk factors for non-Hodgkins lymphoma among men. Occup Environ Med. 2003. 60: E11-
86. Devasahayam G, Burke DJ, Sturgill TW. Golgi manganese transport is required for rapamycin signaling in saccharomyces cerevisiae. Genetics. 2007. 177: 231-8
87. Duncan RJ, Tipton KF. The oxidation and reduction of glyoxylate by lactic dehydrogenase. Eur J Biochem. 1969. 11: 58-61
88. Dupont CL, Tanaka Y. Blood manganese levels in children with convulsive disorder. Biochem Med. 1985. 33: 246-55
89. Dutra BK, Fernandes FA, Failace DM, Oliveira GT. Effect of Roundup (glyphosate formulation) in the energy metabolism and reproductive traits of Hyalella castroi (Crustacea, Amphipoda, Dogielinotidae). Ecotoxicology. 2011. 20: 255-63
90. Dutta U, Cohenford MA, Guha M, Dain JA. Non-enzymatic interactions of glyoxylate with lysine, arginine, and glucosamine: A study of advanced non-enzymatic glycation like compounds. Bioorg Chem. 2007. 35: 11-24
91. Eder W, Ege MJ, von Mutius E. The Asthma Epidemic. N Engl J Med. 2006. 355: 2226-35
92. Eichner K, Reutter M, Wittmann R, Parliament TH, Morello MJ, McGorrin RJ.editors. Detection of Amadori compounds in heated foods. Thermally Generated Flavors (ACS Symposium Series 543). Washington D.C: American Chemical Society; 1994. p.
93. Eisen A, Calne D. Amyotrophic lateral sclerosis, Parkinson'S disease and Alzheimer'S disease: Phylogenetic disorders of the human neocortex sharing many characteristics. Can J Neurol Sci. 1992. 19: 117-23
94. Elder A, Gelein R, Silva V, Feikert T, Opanashuk L, Carter J. Translocation of inhaled ultrafine manganese oxide particles to the central nervous system. Environ Health Perspect. 2006. 114: 1172-8
95. Endo T, Shioiri T, Kitamura H, Kimura T, Endo S, Masuzawa N. Altered chemical metabolites in the amygdala-hippocampus region contribute to autistic symptoms of autism spectrum disorders. Biol Psychiatry. 2007. 62: 1030-7
96. Erickson SK, Lear SR, Deane S, Dubrac S, Huling SL, Nguyen L. Hypercholesterolemia and changes in lipid and bile acid metabolism in male and female cyp7A1-deficient mice. J Lipid Res. 2003. 44: 1001-9
97. Estiasih T, Kgs Ahmadi, Ginting E, Priyanto AD. Modification of soy crude lecithin by partial enzymatic hydrolysis using phosholipase A1. Int Food Res J. 2013. 20: 843-9
98. Ettinger B, Black DM, Mitlak BH, Knickerbocker RK, Nickelsen T, Genant HK. Reduction of vertebral fracture risk in postmenopausal women with osteoporosis treated with raloxifene: Results from a 3-year randomized clinical trial. JAMA. 1999. 282: 637-45
99. Fabiani G, Rogacheski E, Wiederkehr JC, Khouri J, Cianfarano A. Liver transplantion in a patient with rapid onset Parkinsonism-dementia complex induced by manganism secondary to liver failure. Arq Neuropsiquiatr. 2007. 65: 685-8
100. Falchuk KH, Hardy C, Ulpino L, Vallee BL. RNA metabolism, manganese, and RNA polymerases of zinc-sufficient and zinc-deficient Euglena gracilis. Proc Natl Acad Sci. 1978. 75: 4175-9
101. Fallingborg J. Intraluminal pH of the human gastrointestinal tract. Dan Med Bull. 1999. 46: 183-96
102. Federici F, Vitali B, Gotti R, Pasca MR, Gobbi S, Peck AB. Characterization and Heterologous Expression of the Oxalyl Coenzyme A Decarboxylase Gene from Bifidobacterium lactis. Appl Environ Microbiol. 2004. 70: 5066-73
103. Finley JW, Davis CD. Manganese deficiency and toxicity: Are high or low dietary amounts of manganese cause for concern?. BioFactors. 1999. 10: 15-24
104. Fischler B, Cluydts R, De Gucht Y, Kaufman L, De Meirleir K. Generalized anxiety disorder in chronic fatigue syndrome. Acta Psychiatr Scand. 1997. 95: 405-13
105. Flint DH, Tuminello JF, Emptage MH. The inactivation of Fe-S cluster containing hydro-lyases by superoxide. J Biol Chem. 1993. 268: 22369-76
106. Foster JA, Neufeld KA. Gut-brain axis: How the microbiome influences anxiety and depression. Trends Neurosci. 2013. 36: 305-12
107. Fridman O, Goldberg A, Ronin I, Shoresh N, Balaban NQ. Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations. Nature. 2014. 513: 418-21
108. Gasnier C, Dumont C, Benachour N, Clair E, Chagnon MC, Séralini GE. Glyphosate-based herbicides are toxic and endocrine disruptors in human cell lines. Toxicology. 2009. 262: 184-91
109. Ghanizadeh A. Increased glutamate and homocysteine and decreased glutamine levels in autism: A review and strategies for future studies of amino acids in autism. Dis Markers. 2013. 35: 281-6
110. Gillott A. Anxiety in high-functioning children with autism. Autism. 2001. 5: 277-86
111. Giwercman A, Bonde JP. Declining male fertility and environmental factors. Endocrinol Metab Clin North Am. 1998. 27: 807-30
112. Görker I, Tüzün Ü. Autistic-like findings associated with a urea cycle disorder in a 4-year-old girl. Rev Psychiatr Neurosci. 2005. 30: 133-5
113. Gregus Z, Gyurasics A, Csanaky I. Biliary and urinary excretion of inorganic arsenic: Monomethylarsonous acid as a major biliary metabolite in rats. Toxicol Sci. 2000. 56: 18-25
114. Guban J, Korver DR, Allison GE, Tannock GW. Relationship of dietary antimicrobial drug administration with broiler performance, decreased population levels of Lactobacillus salivarius, and reduced bile salt deconjugation in the ileum of broiler chickens. Poult Sci. 2006. 85: 2186-94
115. Gui YX, Fan XN, Wang HM, Wang G, Chen SD. Glyphosate induced cell death through apoptotic and autophagic mechanisms. Neurotoxicol Teratol. 2012. 34: 344-9
116. Guilhermea S, Gaivãob I, Santosa MA, Pachecoa M. DNA damage in fish (Anguilla anguilla) exposed to a glyphosate-based herbicide – Elucidation of organ-specificity and the role of oxidative stress. Mutat Res. 2012. 743: 1-9
117. Guillet G, Poupart J, Basurco J, De Luca V. Expression of tryptophan decarboxylase and tyrosine decarboxylase genes in tobacco results in altered biochemical and physiological phenotypes. Plant Physiol. 2000. 122: 933-43
118. Hancock DA. The feeding behaviour of starfish on Essex oyster beds. J Mar Biol Assoc U.K. 1955. 34: 313-31
119. Harris SR, Feil EJ, Holden MT, Quail MA, Nickerson EK, Chantratita N. Evolution of MRSA during hospital transmission and intercontinental spread. Science. 2010. 327: 469-74
120. Hauser RA, Zesiewicz TA, Rosemurgy AS, Martinez C, Olanow CW. Manganese intoxication and chronic liver failure. Ann Neurol. 1994. 36: 871-5
121. Hawkey PM, Jones AM. The changing epidemiology of resistance. J Antimicrob Chemother. 2009. 64: i3i10-
122. Hawkey PMJones AMLast accessed on 2014 Aug 28. Available from: http://www.epa.gov/pesticides/chemical/foia/cleared-reviews/reviews/103601/103601-2009-02-26a.pdf .
123. Hofmann AF, Eckmann L. How bile acids confer gut mucosal protection against bacteria. Proc Natl Acad Sci U S A. 2006. 103: 4333-4
124. Hollick MF, Chen TC. Vitamin D deficiency a worldwide problem with health consequences. Am J Clin Nutr. 2008. 87: 10805-68
125. Howles PA, Sewalt VJ, Paiva NL, Elkind Y, Bate NJ, Lamb C. Overexpression of L-phenylalanine ammonia-lyase in transgenic tobacco plants reveals control points for flux into phenylpropanoid biosynthesis. Plant Physiol. 1996. 112: 1617-24
126. Huang P, Li G, Chen C, Wang H, Han Y, Zhang S. Differential toxicity of Mn2+and Mn3+to rat liver tissues: Oxidative damage, membrane fluidity and histopathological changes. Exp Toxicol Pathol. 2012. 64: 197-203
127. Huber D. What about glyphosate-induced manganese deficiency?. Fluid J. 2007. p. 20-22
128. Hultberg M. Cysteine turnover in human cell lines is influenced by glyphosate. Environ Toxicol Pharmacol. 2007. 24: 19-22
129. Hurley ES, Woolley DE, Rosenthal F, Timiras PS. Influence of manganese on susceptibility of rats to convulsions. Am J Physiol. 1963. 204: 493-6
130. Ikeda S, Yamaguchi Y, Sera Y, Ohshiro H, Uchino S, Yamashita Y. Manganese deposition in the globus pallidus in patients with biliary atresia. Transplantation. 2000. 69: 2339-43
131. Inoue E, Hori S, Narumi Y, Fujita M, Kuriyama K, Kadota T. Portal-systemic encephalopathy: Presence of basal ganglia lesions with high signal intensity on MR images. Radiology. 1991. 179: 551-5
132. Ito Y, Ishige K, Aizawa M, Fukuda H. Characterization of quinolone antibacterial-induced convulsions and increases in nuclear AP-1 DNA- and CRE-binding activities in mouse brain. Neuropharmacology. 1999. 38: 717-23
133. Jayasumana C, Gunatilake S, Senanayake P. Glyphosate, hard water and nephrotoxic metals: Are they the culprits behind the epidemic of chronic kidney disease of unknown etiology ina Sri Lanka?. Int J Environ Res Public Health. 2014. 11: 2125-47
134. Jiang W, Metcalf WW, Lee KS, Wanner BL. Molecular cloning, mapping, and regulation of Pho regulon genes for phosphonate breakdown by the phosphonatase pathway of Salmonella typhimurium LT2. J Bacteriol. 1995. 177: 6411-21
135. Jiraungkoorskul W, Upatham ES, Kruatrachue M, Sahaphong S, Vichasri-Grams S, Pokethitiyook P. Histopathological effects of Roundup, a glyphosate herbicide, on Nile tilapia (Oreochromis niloticus). Sci Asia. 2002. 28: 121-7
136. Joyce SA, MacSharry J, Casey PG, Kinsell M, Murphy EF, Shanahan F. Regulation of host weight gain and lipid metabolism by bacterial bile acid modification in the gut. Proc Natl Assoc Sci U S A. 2014. 111: 7421-6
137. Kakulavarapu V, Rao R, Norenberg MD. Manganese induces the mitochondrial permeability transition in cultured astrocytes. J Biol Chem. 2004. 279: 32333-8
138. Kaneta H, Fujii J, Suzuki K, Kasai H, Kawamore R, Kamadaa T. DNA cleavage induced by glycation of Cu, Zn-superoxide dismutase. Biochem J. 1994. 304: 219-25
139. Kannus P, Palvanen M, Niemi S, Parkkari J, Järvinen M, Ilkka Vuori I. Osteoporotic fractures of the proximal humerus in elderly Finnish persons: Sharp increase in 1970-1998 and alarming projections for the new millennium. Acta Orthop Scand. 2000. 71: 465-70
140. Kannus P, Parkkari J, Sievnen H, Heinonen A, Vuori I, Järvinen M. Epidemiology of hip fractures. Bone. 1996. 18: 557-63
141. Kaplan M, Rubaltelli FF, Hammerman C, Vilei MT, Leiter C, Abramov A. Conjugated bilirubin in neonates with glucose-6-phosphate dehydrogenase deficiency. J Pediatr. 1996. 128: 695-7
142. Kaplan MRubaltelli FFHammerman CVilei MTLeiter CAbramov ALast accessed on 2014 Sep 15. Available from: http://www.google.com/patents/US5464806 .
143. Kearney PC, Kaufman DD.editors. Herbicides Chemistry: Degradation and Mode of Action. USA: CRC Press; 1988. p.
144. Kehres DG, Maguire ME. Emerging themes in manganese transport, biochemistry and pathogenesis in bacteria. FEMS Microbiol Rev. 2003. 27: 263-90
145. Keller KA, Barnes PD. Rickets vs abuse: A national and international epidemic. Pediatr Radiol. 2008. 38: 1210-6
146. Kiely T, Donaldson D, Grube A.editors. Pesticides industry sales and usage-2000 and 2001 market estimates. Washington, DC: US Environmental Protection Agency; 2004. p.
147. Kim YH, Hong JR, Gil HW, Song HY, Hong SY. Mixtures of glyphosate and surfactant TN20 accelerate cell death via mitochondrial damage-induced apoptosis and necrosis. Toxicol In Vitro. 2013. 27: 191-7
148. Kirschbaum BB, Schoolwerth AC. Acute aluminum toxicity associated with oral citrate andaluminum-containing antacids. Am J Med Sci. 1989. 297: 9-11
149. Klevay LM. Alzheimer'S disease as copper deficiency. Med Hypotheses. 2008. 70: 802-7
150. Klos KJ, Ahlskog JE, Josephs KA, Fealey RD, Cowl CT, Kumar N. Neurologic spectrum of chronic liver failure and basal ganglia T1 hyperintensity on magnetic resonance imaging: Probable manganese neurotoxicity. Arch Neurol. 2005. 62: 1385-90
151. Kohanski MA, DePristo MA, Collins JJ. Sublethal antibiotic treatment leads to multidrug resistance via radical-induced mutagenesis. Mol Cell. 2010. 37: 311-20
152. Konstantynowicz J, Porowski T, Zoch-Zwierz W, Wasilewska J, Kadziela-Olech H, Kulak W. A potential pathogenic role of oxalate in autism. Eur J Paediatr Neurol. 2012. 16: 485-91
153. Krüger M, Schledorn P, Schrödl W, Hoppe HW, Lutz W, Shehata AA. Detection of glyphosate residues in animals and humans. J Environ Anal Toxicol. 2014. 4: 2-
154. Krüger M, Schrödl W, Neuhaus J, Shehata AA. Field investigations of glyphosate in urine of Danish dairy cows. J Environ Anal Toxicol. 2013. 3: 1-7
155. Krüger M, Schrödl W, Pedersen I, Shehata AA. Detection of glyphosate in malformed piglets. J Environ Anal Toxicol. 2014. 4: 5-
156. Larsen K, Najle R, Lifschitz A, Maté ML, Lanusse C, Virkel GL. Effects of sublethal exposure to a glyphosate-based herbicide formulation on metabolic activities of different xenobiotic-metabolizing enzymes in rats. Int J Toxicol. 2014. 33: 307-18
157. Leach RM. Role of manganese in the synthesis of mucopolysaccharides. Fed Proc. 1967. 26: 118-20
158. Leach RM, Muenster AM, Wien EM. Studies on the role of manganese in bone formation: II. Effect upon chondroitin sulfate synthesis in chick epiphyseal cartilage. Arch Biochem Biophys. 1969. 133: 22-8
159. Lebovitz RM, Zhang H, Vogel H, Cartwright J, Dionne L, Lu N. Neurodegeneration, myocardial injury, and perinatal death in mitochondrial superoxide dismutase-deficient mice. Proc Natl Acad Sci USA. 1996. 93: 9782-7
160. Leffler CT, Philippi AF, Leffler SG, Mosure JC, Kim PD. Glucosamine, chondroitin, and manganese ascorbate for degenerative joint disease of the knee or low back: A randomized, double-blind, placebo-controlled pilot study. Mil Med. 1999. 164: 85-91
161. Levin J, Bertsch U, Kretzschmar H, Giese A. Single particle analysis of manganese-induced prion protein aggregates. Biochem Biophys Res Commun. 2005. 329: 1200-7
162. Levisohn PM. The autism-epilepsy connection. Epilepsia. 2007. 48: 33-5
163. Levy BS, Nassetta WJ. Neurologic effects of manganese in humans: A review. Int J Occup Environ Health. 2003. 9: 153-63
164. Li Y, Huang TT, Carlson EJ, Melov S, Ursell PC, Olson JL. Dilated cardiomyopathy and neonatal lethality in mutant mice lacking manganese superoxide dismutase. Nat Genet. 1995. 11: 376-81
165. Lin Q. Submerged fermentation of Lactobacillus rhamnosus YS9 for γ-aminobutyric acid (GABA) production. Braz J Microbiol. 2013. 44: 183-7
166. Lia C, Nie SP, Ding Q, Zhu KX, Wang ZJ, Xiong T. Cholesterol-lowering effect of Lactobacillus plantarum NCU116 in a hyperlipidaemic rat model. J Funct Foods. 2014. 8: 340-7
167. Lin J, Lee IS, Frey J, Slonczewski JL, Foster JW. Comparative analysis of extreme acid survival in Salmonella typhimurium, Shigella flexneri, and Escherichia coli. J Bacteriol. 1995. 177: 4097-104
168. Lindquist NG, Larsson BS, Lydén-Sokolowski A. Neuromelanin and its possible protective and destructive properties. Pigment Cell Res. 1987. 1: 133-6
169. Livermore DM. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare?. Clin Infect Dis. 2002. 34: 634-40
170. Lopes FM, Varela Junior AS, Corcini CD, da Silva AC, Guazzelli VG, Tavares G. Effect of glyphosate on the sperm quality of zebrafish Danio rerio. Aquat Toxicol. 2014. 155: 322-6
171. Lu W, Li L, Chen M, Zhou Z, Zhang W, Ping S. Genome-wide transcriptional responses of Escherichia coli to glyphosate, a potent inhibitor of the shikimate pathway enzyme 5-enolpyruvylshikimate-3-phosphate synthase. Mol Biosyst. 2013. 9: 522-30
172. Lucchini RG, Martin CJ, Doney BC. From manganism to manganese-induced Parkinsonism: A conceptual model based on the evolution of exposure. Neuromol Med. 2009. 11: 311-21
173. Lundager Madsen HE, Christensen HH, Gottlieb-Petersen C, Andresen AF, Smidsrod O, Pontchour CO. Stability Constants of Copper (II), Zinc, Manganese (II), calcium, and magnesium complexes of N-(phosphonomethyl) glycine (Glyphosate). Acta Chem Scand A. 1978. 32: 79-83
174. Lushchak OV, Kubrak OI, Storey JM, Storey KB, Lushchak VI. Low toxic herbicide Roundup induces mild oxidative stress in goldfish tissues. Chemosphere. 2009. 76: 932-7
175. Lyke K. A study on the effects of a high soy-content diet on urinary oxalate levels in humans. PhD Thesis, Hawthorn University Whitethorn, CA. January. 2013. p.
176. MacDonald MG. Hidden risks: Early discharge and bilirubin toxicity due to glucose 6-phosphate dehydrogenase deficiency. Pediatrics. 1995. 96: 734-8
177. Maes M, Mihaylova I, Leunis JC. Increased serum IgA and IgM against LPS of enterobacteria in chronic fatigue syndrome (CFS): Indication for the involvement of gram-negative enterobacteria in the etiology of CFS and for the presence of an increased gut intestinal permeability. J Affect Disord. 2007. 99: 237-40
178. Magnus Ø, Brekke I, Åbyholm T, Purvis K. Effects of manganese and other divalent cations on progressive motility of human sperm. Syst Biol Reprod Med. 1990. 25: 159-66
179. Malecki EA, Radzanowski GM, Radzahowski TJ, Gallaher DD, Greger JL. Biliary manganese excretion in conscious rats is affected by acute and chronic manganese intake but not by dietary fat. J Nutr. 1996. 126: 489-98
180. Marin JJ. Bile acids: Chemistry, physiology, and pathophysiology. World J Gastroenterol. 2009. 15: 804-16
181. Masánová V, Mitrova E, Ursinyova M, Uhnakova I, Slivarichova D. Manganese and copper imbalance in the food chain constituents in relation to Creutzfeldt-Jakob disease. Int J Environ Health Res. 2007. 17: 419-28
182. Matés JM, Sánchez-Jiménez F. Antioxidant enzymes and their implications in pathophysiologic processes. Front Biosci. 1999. 4: d339-45
183. McDonald LC, Killgore GE, Thompson A, Owens RC, Kazakova SV, Sambol SP. An epidemic, toxin genevariant strain of Clostridium difficile. N Engl J Med. 2005. 353: 2433-41
184. McLaren PJ, Cave JG, Parker EM, Slocombe RF. Chondrodysplastic calves in Northeast Victoria. Vet Pathol. 2007. 44: 342-54
185. Melø TM, Larsen C, White LR, Aasly J, Sjøbakk TE, Flaten TP. Manganese, copper, and zinc in cerebrospinal fluid from patients with multiple sclerosis. Biol Trace Elem Res. 2003. 93: 1-8
186. Melov S, Schneider JA, Day BJ, Hinerfeld D, Coskun P, Mirra SS. A novel neurological phenotype in mice lacking mitochondrial manganese superoxide dismutase. Nat Genet. 1998. 18: 159-63
187. Menendez A, Arena ET, Guttman JA, Thorson L, Vallance BA, Vogl W. Salmonella infection of gallbladder epithelial cells drives local inflammation and injury in a model of acute typhoid fever. J Infect Dis. 2009. 200: 1703-13
188. Mesnage R, Bernay B, Séralini GE. Ethoxylated adjuvants of glyphosate-based herbicides are active principles of human cell toxicity. Toxicology. 2013. 313: 122-8
189. Mesnage R, Defarge N, de Vendômois JS, Séralini GE. Major pesticides are more toxic to human cells than their declared active principles. Biomed Res Int 2014. 2014. p.
190. Micek ST, Lloyd AE, Ritchie DJ, Reichley RM, Fraser VJ, Kollef MH. Pseudomonas aeruginosa bloodstream infection: Importance of appropriate initial antimicrobial treatment. Antimicrob Agents Chemother. 2005. 49: 1306-11
191. Molbak K, Baggesen DL, Aarestrup FM, Ebbesen JM, Engberg J, Frydendahl K. An outbreak of multidrug-resistant, quinolone-resistant Salmonella enterica serotype typhimurium DT104. N Engl J Med. 1999. 341: 1420-5
192. Moore JK, Braymer HD, Larson AD. Isolation of a Pseudomonas sp. which utilizes the phosphonate herbicide glyphosate. Appl Environ Microbiol. 1983. 46: 316-20
193. Morley WA, Seneff S. Diminished brain resilience: A modern day neurological pathology of increased susceptibility to mild brain trauma, concussion and downstream neurodegeneration. Surg Neurol Int. 2014. 5: 97-
194. Motekaitis RJ, Martell AE. Metal chelate formation by N-Phosphono-methylglycine and related ligands. J Coord Chem. 1985. 14: 139-49
195. Murri M, Leiva I, Gamez-Zumaquero JM, Tinahones FJ, Cardona F, Soriguer F. Gut microbiota in children with type 1 diabetes differs from that in healthy children: A case-control study. BMC Med. 2013. 11: 46-
196. Myhill S, Booth NE, McLaren-Howard J. Chronic fatigue syndrome and mitochondrial dysfunction. Int J Clin Exp Med. 2009. 2: 1-16
197. Napoli E, Wong S, Giulivi C. Evidence of reactive oxygen species-mediated damage to mitochondrial DNA in children with typical autism. Mol Autism. 2013. 4: 2-
198. Napoli E, Wong S, Hertz-Picciotto I, Giulivi C. Deficits in bioenergetics and impaired immune response in granulocytes from children with autism. Pediatrics. 2014. 133: e1405-10
199. Narayan KM, Gregg EW, Fagot-Campagna A, Engelgau MM, Vinicor F. Diabetes a common, growing, serious, costly, and potentially preventable public health problem. Diabetes Res Clin Pract. 2000. 50: S77-84
200. Nebert DW. Clinical importance of the cytochromes P450. Lancet. 2002. 360: 1155-62
201. Negga R, Stuart JA, Machen ML, Salva J, Lizek AJ, Richardson SJ. Exposure to glyphosate- and/or Mn/Zn-ethylene-bisdithiocarbamate-containing pesticides leads to degeneration of γ-aminobutyric acid and dopamine neurons in Caenorhabditis elegans. Neurotox Res. 2012. 21: 281-90
202. Nevison CD. A comparison of temporal trends in United States autism prevalence to trends in suspected environmental factors. Environ Health. 2014. 13: 73-
203. Nishida T, Gaitmatan Z, Che M, Arias IM. Rat liver canalicular membrane vesicles contain an ATP-dependent bile acid transport system. Proc Natl Acad Sci U S A. 1991. 88: 6590-4
204. Norenberg MD. The distribution of glutamine synthetase in the central nervous system. J Histochem Cytochem. 1979. 27: 469-75
205. Norlin M, Wikvall K. Enzymes in the conversion of cholesterol into bile acids. Curr Mol Med. 2007. 7: 199-218
206. Nowell LH, Capel PD, Dileanis PD.editors. Pesticides in stream sediment and aquatic biota- distribution, trends, and governing factors. In: Pesticides in the Hydrologic System series. Boca Raton, Florida: CRC Press; 1999. 4: 1040-
207. Oelzner P, Müller A, Deschner F, Hüller M, Abendroth K, Hein G. Relationship between disease activity and serum levels of vitamin D metabolites and PTH in rheumatoid arthritis. Calcif Tissue Int. 1998. 62: 193-8
208. Okonkwo FO, Ejike CE, Anoka AN, Onwurah IN. Toxicological studies on the short term exposure of Clarias albopunctatus (Lamonte and Nichole 1927) to sub-lethal concentrations of Roundup. Pak J Biol Sci. 2013. 16: 939-44
209. Opresko LK, Brokaw CJ. cAMP-dependent phosphorylation associated with activation of motility of Ciona sperm flagella. Gamete Res. 1983. 8: 201-18
210. O’Rand MG, Widgren EE, Beyler S, Richardson RT. Inhibition of human sperm motility by contraceptive anti-eppin antibodies from infertile male monkeys: Effect on cyclic adenosine monophosphate. Biol Reprod. 2009. 80: 279-85
211. Padda MS, Sanchez M, Akhtar AJ, Boyer JL. Drug induced cholestasis. Hepatology. 2011. 53: 1377-87
212. Paganelli A, Gnazzo V, Acosta H, López SL, Carrasco AE. Glyphosate-based herbicides produce teratogenic effects on vertebrates by impairing retinoic acid signaling. Chem Res Toxicol. 2010. 23: 1586-95
213. Park LC, Albers DS, Xu H, Lindsay JG, Beal MF, Gibson GE. Mitochondrial impairment in the cerebellum of the patients with progressive supranuclear palsy. J Neurosci Res. 2001. 66: 1028-34
214. Park LCAlbers DSXu HLindsay JGBeal MFGibson GELast accessed on 2014 Sep 04. Available from: http://www.mass.gov/eohhs/gov/newsroom/press-releases/dph/closure-of-oyster-beds-in-edgartown-announced.html .
215. Peixoto F. Comparative effects of the Roundup and glyphosate on mitochondrial oxidative phosphorylation. Chemosphere. 2005. 61: 1115-22
216. Perl DP, Warren OC. The neuropathology of manganese-induced Parkinsonism. J Neuropathol Exp Neurol. 2007. 66: 675-82
217. Pikuleva IA. Cholesterol-metabolizing cytochromes P450. Drug Metab Dispos. 2006. 34: 513-20
218. Polak T, Weise D, Metzger F, Ehlis AC, Langer JB, Schramm A. Vagus nerve somatosensory evoked potentials in Parkinsons disease. J Neurol. 2011. 258: 2276-7
219. Porter JW, Dustan P, Jaap WC, Patterson KL, Kosmynin V, Meier OW. Patterns of spread of coral disease in the Florida Keys. Hydrobiologia. 2001. 460: 1-24
220. Post SM, Duez H, Gervois PP, Staels B, Kuipers F, Princen HM. Fibrates Suppress Bile Acid Synthesis via Peroxisome Proliferatoractivated receptor-αmediated downregulation of cholesterol 7α-hydroxylase and sterol 27-hydroxylase expression. Arterioscler Thromb Vasc Biol. 2001. 21: 1840-5
221. Powles SB. Evolved glyphosate-resistant weeds around the world: Lessons to be learnt. Pest Manag Sci. 2008. 64: 360-5
222. Prouty AM, Schwesinger WH, Gunn JS. Biofilm formation and interaction with the surfaces of gallstones by Salmonella spp. Infect Immun. 2002. 70: 2640-9
223. Prusiner SB. Molecular biology of prion disease. Science. 1991. 252: 1515-22
224. Prusiner SB. Biology and genetics of prions causing neurodegeneration. Annu Rev Genet. 2013. 47: 601-23
225. Pullinger CR, Eng C, Salen G, Shefer S, Batta AK, Erickson SK. Human cholesterol 7α-hydroxylase (CYP7A1) deficiency has a hypercholesterolemic phenotype. J Clin Invest. 2002. 110: 109-17
226. Purcell AE, Jeon OH, Zimmerman AW, Blue ME, Pevsner J. Postmortem brain abnormalities of the glutamate neurotransmitter system in autism. Neurology. 2001. 57: 1618-28
227. Purdey JM. Ecosystems supporting clusters of sporadic TSEs demonstrate excesses of the radical-generating divalent cation manganese and deficiencies of antioxidant co factors Cu, Se, Fe, Zn. Med Hypotheses. 2000. 54: 278-306
228. Purdey JM. Elevated levels of ferromagnetic metals in food chains supporting the Guam cluster of neurodegeneration: Do metal nucleated crystal contaminants evoke magnetic fields that initiate the progressive pathogenesis of neurodegeneration?. Med Hypotheses. 2004. 63: 793-809
229. Purdey JM. Metal microcrystal pollutants: The heat resistant, transmissible nucleating agents that initiate the pathogenesis of TSEs?. Med Hypotheses. 2005. 65: 448-77
230. Purgel M, Takács Z, Jonsson CM, Nagy L, Andersson I, Bányai I. Glyphosate complexation to aluminium (III). An equilibrium and structural study in solution using potentiometry, multinuclear NMR, ATRFTIR, ESI-MS and DFT calculations. J Inorg Biochem. 2009. 103: 1426-38
231. Rallis C, Bähler J. Inhibition of TORC1 signaling and increased lifespan: Gained in translation?. Aging. 2013. 5: 335-6
232. Rao AV, Bested AC, Beaulne TM, Katzman MA, Iorio C, Berardi JM. A randomized, double-blind, placebo-controlled pilot study of a probiotic in emotional symptoms of chronic fatigue syndrome. Gut Pathog. 2009. 1: 6-
233. Ramachandra R, Namburi RB, Ortega-Martinez O, Shi X, Zaia J, Dupont ST. Brittlestars contain highly sulfated chondroitin sulfates/dermatan sulfates that promote fibroblast growth factor 2-induced cell signaling. Glycobiology. 2014. 24: 195-207
234. Reitsma S, Slaaf DW, Vink H, van Zandvoort MA, oude Egbrink MG. The endothelial glycocalyx: Composition, functions, and visualization. Pflugers Arch. 2007. 454: 345-59
235. Richard IH. Anxiety disorders in Parkinson'S disease. Adv Neurol. 2005. 96: 42-55
236. Rodionov DA, Vitreschak AG, Mironov AA, Gelfand MS. Comparative genomics of the vitamin B12 metabolism and regulation in prokaryotes. J Biol Chem. 2003. 278: 41148-59
237. Rolo AP, Oliveira PJ, Moreno AJM, Palmeira CM. Bile acids affect liver mitochondrial bioenergetics: Possible relevance for cholestasis therapy. Toxicol Sci. 2000. 57: 177-85
238. Román GC, Ghassabian A, Bongers-Schokking JJ, Jaddoe VW, Hofman A, de Rijke YB. Association of gestational maternal hypothyroxinemia and increased autism risk. Ann Neurol. 2013. 74: 733-42
239. Rose S, Melnyk S, Pavliv O, Bai S, Nick TG, Frye RE. Evidence of oxidative damage and inflammation associated with low glutathione redox status in the autism brain. Transl Psychiatry. 2012. 2: e134-
240. Rossi M, Amaretti A, Raimondi S. Folate production by probiotic bacteria. Nutrients. 2011. 3: 118-34
241. Rossignol DA, Frye RE. Mitochondrial dysfunction in autism spectrum disorders: A systematic review and meta-analysis. Mol Psychiatry. 2012. 17: 290-314
242. Rossignol DA, Genuis SJ, Frye RE. Environmental toxicants and autism spectrum disorders: A systematic review. Transl Psychiatry. 2014. 4: e360-
243. Rossignol DA, Frye RE. Evidence linking oxidative stress, mitochondrial dysfunction, and inflammation in the brain of individuals with autism. Front Physiol. 2014. 22: 150-
244. Roth JA. Homeostatic and toxic mechanisms regulating manganese uptake, retention, and elimination. Biol Res. 2006. 39: 45-57
245. Rotig A, de Lonlay P, Chretien D, Foury F, Koenig M, Sidi D. Aconitase and mitochondrial iron-sulphur protein deficiency in Friedreich ataxia. Nat Genet. 1997. 17: 215-7
246. Rowe B, Ward LR, Threlfall EJ. Multidrug-resistant Salmonella typhi: A worldwide epidemic. Clin Infect Dis. 1997. 24: S106-9
247. Saltman PD, Strause LG. The role of trace minerals in osteoporosis. J Am Coll Nutr. 1993. 12: 384-9
248. Samsel A, Seneff S. Glyphosate'S suppression of cytochrome P450 enzymes and amino acid biosynthesis by the gut microbiome: Pathways to modern diseases. Entropy. 2013. 15: 1416-63
249. Samsel A, Seneff S. Glyphosate, pathways to modern diseases II: Celiac sprue and gluten intolerance. Interdiscip Toxicol. 2013. 6: 159-84
250. Scales CD, Smith AC, Hanley JM, Saigal CS. Prevalence of kidney stones in the United States. Eur Urol. 2012. 62: 160-5
251. Schapira AH. Mitochondria in the aetiology and pathogenesis of Parkinsons disease. Lancet Neurol. 2008. 7: 97-109
252. Schnedler N, Burckhardt G, Burckhardt BC. Glyoxylate is a substrate of the sulfate-oxalate exchanger, SAT-1, and increases its expression in HepG2 cells. J Hepatol. 2011. 54: 513-20
253. Schinasi L, Leon ME. Non-Hodgkin lymphoma and occupational exposure to agricultural pesticide chemical groups and active ingredients: A systematic review and meta-analysis. Int J Environ Res Public Health. 2014. 11: 4449-527
254. Shrader RE, Erway LC, Hurley LS. Mucopolysaccharide synthesis in the developing inner ear of manganese-deficient and pallid mutant mice. Teratology. 1973. 8: 257-66
255. Shrader REErway LCHurley LSLast accessed on 2014 Sep 10. 2321.
256. Seiler N. Ammonia and Alzheimers disease. Neurochem Int. 2002. 41: 189-207
257. Selvam R, Bijikurien T. Methionine feeding prevents kidney stone deposition by restoration of free radical mediated changes in experimental urolithiasis. J Nutr Biochem. 1991. 2: 644-51
258. Selvam R. Calcium oxalate stone disease: Role of lipid peroxidation and antioxidants. Urol Res. 2002. 30: 35-47
259. Selvan T, Rajiah K, Nainar MS, Mathew EM. A clinical study on glucosamine sulfate versus combination of glucosamine sulfate and NSAIDs in mild to moderate knee osteoarthritis. ScientificWorldJournal 2012. 2012. p.
260. Seneff S, Lauritzen A, Davidson RM, Lentz-Marino L. Is encephalopathy a mechanism to renew sulfate in autism?. Entropy. 2013. 15: 372-406
261. Séralini GE, Clair E, Mesnage R, Gress S, Defarge N, Malatesta M. Republished study: Long-term toxicity of a Roundup herbicide and a Roundup-tolerant genetically modified maize. Environ Sci Eur. 2014. 26: 14-
262. Sharma AK, Gaur K, Tiwari RK, Gaur MS. Computational interaction analysis of organophosphorus pesticides with different metabolic proteins in humans. J Biomed Res. 2011. 25: 335-47
263. Shehata AA, Schrödl W, Aldin AA, Hafez HM, Krüger M. The effect of glyphosate on potential pathogens and beneficial members of poultry microbiota in vitro. Curr Microbiol. 2013. 66: 350-8
264. Shehata AA, Schrödl W, Schledorn P, Krüger M. Distribution of glyphosate in chicken organs and its reduction by humic acid supplementation. J Poult Sci. 2014. 51: 333-
265. Shimmura C, Suda S, Tsuchiya KJ, Hashimoto K, Ohno K, Matsuzaki H. Alteration of plasma glutamate and glutamine levels in children with high-functioning autism. PLoS ONE. 2011. 6: e25340-
266. Shipley MT, Halloran FJ, de la Torre J. Surprisingly rich projection from locus coeruleus to the olfactory bulb in the rat. Brain Res. 1985. 329: 294-9
267. Sidoryk-Wegrzynowicz M, Aschner M. Manganese toxicity in the central nervous system: The glutamine/glutamate-c-aminobutyric acid cycle. J Intern Med. 2013. 273: 466-77
268. Soderland P, Lovekar S, Weiner DE, Brooks DR, Kaufman JS. Chronic kidney disease associated with environmental toxins and exposures. Adv Chronic Kidney Dis. 2010. 17: 254-64
269. Sohler A, Pfeiffer CC. A direct method for the determination of manganese in whole blood. Patients with seizure activity have low blood levels. J Orthomol Psych. 1979. 8: 275-80
270. Soldin OP, Aschner M. Effects of manganese on thyroid hormone. Neurotoxicology. 2007. 28: 951-6
271. Ståhlberg D, Reihnér E, Rudling M, Berglund L, Einarsson K, Angelin B. Influence of bezafibrate on hepatic cholesterol metabolism in gallstone patients: Reduced activity of cholesterol 7alpha-hydroxylase. Hepatology. 1995. 21: 1025-30
272. Stevens DJ, Walter ED, Rodriguez A, Draper D, Davies P, Brown DR. Early onset prion disease from octarepeat expansion correlates with copper binding properties. PLoS Pathogens. 2009. 5: e1000390-
273. Stieger B. Regulation of the expression of the hepatocellular sulfate-oxalate exchanger SAT-1 (SLC26A1) by glyoxylate: A metabolic link between liver and kidney?. J Hepatol. 2011. 54: 406-7
274. Stoyanovsky DA, Huang Z, Jiang J, Belikova NA, Tyurin V, Epperly MW. A manganeseporphyrin complex decomposes H2O2, inhibits apoptosis, and acts as a radiation mitigator in vivo. ACS Med Chem Lett. 2011. 2: 814-7
275. Strange RC, Spiteri MA, Ramachandran S, Fryer AA. Glutathione-S-transferase family of enzymes. Mutat Res. 2001. 482: 21-6
276. Strause LG, Hegenauer J, Saltman P, Cone R, Resnick D. Effects of long-term dietary manganese and copper deficiency on rat skeleton. J Nutr. 1986. 116: 135-41
277. Streifel KM, Miller J, Mouneimne R, Tjalkens RB. Manganese inhibits ATP-induced calcium entry through the transient receptor potential channel TRPC3 in astrocytes. Neurotoxicology. 2013. 34: 160-6
278. Strople J, Lovell G, Heubi J. Prevalence of subclinical vitamin K deficiency in cholestatic liver disease. J Pediatr Gastroenterol Nutr. 2009. 49: 78-84
279. Subramanian N, Qadri A. Lysophospholipid sensing triggers secretion of flagellin from pathogenic salmonella. Nat Immunol. 2006. 7: 583-9
280. Svedruzić D, Jónsson S, Toyota CG, Reinhardt LA, Ricagno S, Lindqvist Y. The enzymes of oxalate metabolism: Unexpected structures and mechanisms. Arch Biochem Biophys. 2005. 433: 176-92
281. Swerdlow RH, Burns JM, Khan SM. The Alzheimer'S disease mitochondrial cascade hypothesis. J Alzheimers Dis. 2010. 20: 265-79
282. Tabrizi SJ, Workman J, Hart PE, Mangiarini L, Mahal A, Bates G. Mitochondrial dysfunction and free radical damage in the Huntington R6/2 transgenic mouse. Ann Neurol. 2000. 47: 80-6
283. Takeda A, Kodama Y, Ishiwatari S, Okada S. Manganese transport in the neural circuit of rat CNS. Brain Res Bull. 1998. 45: 149-52
284. Takeda A, Sawashita J, Okada S. Manganese uptake into rat brain during development and aging. J Neurosci Res. 1999. 56: 93-8
285. Takeuchi H, Jin S, Wang J, Zhang G, Kawanokuchi J, Kuno R. Tumor necrosis factor-alpha induces neurotoxicity via glutamate release from hemichannels of activated microglia in an autocrine manner. J Biol Chem. 2006. 281: 21362-8
286. Tannock GW, Wood BJ.editors. Lactic microbiota of pigs, mice and rats. The lactic acid bacteria in health and disease. London, United Kingdom: Elsevier Applied Science; 1992. 1:
287. Tash JS, Hidaka H, Means AR. Axokinin phosphorylation by cAMP-dependent protein kinase is sufficient for activation of sperm flagellar motility. J Cell Biol. 1986. 103: 649-55
288. Teri L, Ferretti LE, Gibbons LE, Logsdon RG, McCurry SM, Kukull WA. Anxiety in Alzheimer'S disease: Prevalence and comorbidity. J Gerontol A Biol Sci Med Sci. 1999. 54: M348-52
289. Thongprakaisang S, Thiantanawat A, Rangkadilok N, Suriyo T, Satayavivad J. Glyphosate induces human breast cancer cells growth via estrogen receptors. Food Chem Toxicol. 2013. 59: 129-36
290. Thrane VR, Thrane AS, Wang F, Cotrina ML, Smith NA, Chen M. Ammonia triggers neuronal disinhibition and seizures by impairing astrocyte potassium buffering. Nat Med. 2013. 19: 1643-8
291. Tjälve H, Henriksson J, Tallkvist J, Larsson BS, Lindquist NG. Uptake of manganese and cadmium from the nasal mucosa into the central nervous system via olfactory pathways in rats. Pharmacol Toxicol. 1996. 79: 347-56
292. Tsolis RM, Baumler AJ, Heffron F. Role of Salmonella typhimurium Mn-superoxide dismutase (SodA) in protection against early killing by J774 macrophages. Infect Immun. 1995. 63: 1739-44
293. Tuchman R, Rapin I. Epilepsy in autism. Lancet Neurol. 2002. 1: 352-8
294. Tzin V, Galili G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol Plant. 2010. 3: 956-72
295. Ullah N, Khan FM, Mukhtiar M. Human blood glutathione (GSH) as a tool for arsenic detoxification. Afr J Pharm Pharmacol. 2012. 6: 17-23
296. Undabeytia T, Morillo E, Maqueda C. FTIR study of glyphosate-copper complexes. J Agric Food Chem. 2002. 50: 1918-21
297. Undabeytia TMorillo EMaqueda CLast accessed on 2014 May 14. Available from: http://www.ams.usda.gov/pdp .
298. Undabeytia TMorillo EMaqueda CLast accessed on 2014 Jun 30. Available from: http://www.google.com/patents/US7771736 .
299. Undabeytia TMorillo EMaqueda CLast accessed on 2014 Jun 30. Available from: http://www.google.com/patents/US8507409 .
300. Vielhaber S, Niessen HG, Debska-Vielhaber G, Kudin AP, Wellmer J, Kaufmann J. Subfield-specific loss of hippocampal N-acetyl aspartate in temporal lobe epilepsy. Epilepsia. 2008. 49: 40-50
301. Visner GA, Dougall WC, Wilson JM, Burr IA, Nick HS. Regulation of manganese superoxide dismutase by lipopolysaccharide, interleukin-1, and tumor necrosis factor: Role in the acute inflammatory response. J Biol Chem. 1990. 265: 2856-64
302. Volkmar FR, Nelson DS. Seizure disorders in autism. J Am Acad Child Adolesc Psychiatry. 1990. 29: 127-9
303. Wandernoth PM, Raubuch M, Mannowetz N, Becker HM, Deitmer JW, Sly WS. Role of carbonic anhydrase IV in the bicarbonate-mediated activation of murine and human sperm. PLoS ONE. 2010. 5: e15061-
304. Wang G, Fan XN, Tan YY, Cheng Q, Chen SD. Parkinsonism after chronic occupational exposure to glyphosate. Parkinsonism Relat Disord. 2011. 17: 486-7
305. Watts RJ, Sarasa J, Loge FJ, Teel AL. Oxidative and reductive pathways in manganese-catalyzed Fentons reactions. J Environ Eng. 2005. 131: 158-64
306. Weinstein DL, Carsiotis M, Lissne CR, O’Brien AD. Flagella help Salmonella ty- phimurium survive within murine macrophages. Infect Immun. 1984. 46: 819-25
307. Weinstein DLCarsiotis MLissne CRO’Brien AD. Available from: http://www.usatoday.com/story/news/nation/2013/12/28/starfish-dying-wasting-disease-mysery/4208859/ .
308. Wennberg RP. The blood-brain barrier and bilirubin encephalopathy. Cell Mol Neurobiol. 2000. 20: 97-109
309. Werner P, Pitt D, Raine CS. Multiple sclerosis: Altered glutamate homeostasis in lesions correlates with oligodendrocyte and axonal damage. Ann Neurol. 2001. 50: 169-80
310. Westmark CJ. Soy infant formula and seizures in children with autism: A retrospective study. PLoS ONE. 2014. 9: e80488-
311. Wichmann F, Udikovic-Kolic N, Andrew S, Handelsmana J. Diverse antibiotic resistance genes in dairy cow manure. MBio. 2014. 5: e01017-
312. Willis AW, Evanoff BA, Lian M, Galarza A, Wegrzyn A, Schootman M. Metal emissions and urban incident Parkinson disease: A community health study of medicare beneficiaries by using geographic information systems. Am J Epidemiol. 2010. 172: 1357-63
313. Winter SE, Thiennimitr P, Winter MG, Butler BP, Huseby DL, Crawford RW. Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature. 2010. 467: 426-9
314. Woodburn AT. Glyphosate: Production, pricing and use worldwide. Pest Manag Sci. 2000. 56: 309-12
315. Xu XC, Brinker RJ, Reynolds TL, Abraham W, Graham JA. Pesticide compositions containing oxalic acid. US Patent number 6,992,045 B2, Monsanto Technology LLC, Jan.31. 2006. p.
316. Yao J, Irwin RW, Zhao L, Nilsen J, Hamilton RT, Brinton RD. Mitochondrial bioenergetic deficit precedes Alzheimer'S pathology in female mouse model of Alzheimer'S disease. Proc Natl Acad Sci USA. 2009. 106: 14670-5
317. Yang H, Ramani K, Xia M, Ko KS, Li TW, Oh P. Dysregulation of glutathione synthesis during cholestasis in mice: Molecular mechanisms and therapeutic implications. Hepatology. 2009. 49: 1982-91
318. Yokota T, Gots JS. Requirement of adenosine 3’,5’- cyclicphosphate for flagella formation in Escherichia coli and Salmonella typhimurium. J Bacteriol. 1970. 103: 513-6
319. Zaidi A, Khan MS, Rizvi PQ. Effect of herbicides on growth, nodulation and nitrogen content of greengram. Agron Sustain Dev. 2005. 25: 497-504
320. Ziomber A, Thor P, Krygowska-Wajs A, Załęcki T, Moskała M, Romańska I. Chronic impairment of the vagus nerve function leads to inhibition of dopamine but not serotonin neurons in rat brain structures. Pharmacol Rep. 2012. 64: 1359-67
321. Antoniou M, Habib ME, Howard CV, Jennings RC, Leifert C, Nodari RO. Teratogenic Effects of Glyphosate-Based Herbicides: Divergence of Regulatory Decisions from Scientific Evidence. J Environ Anal Toxicol. 2012. S4: 006-
322. Boocock MR, Coggins JR. Kinetics of 5-enolpyruvylshikimate-3-phosphate synthase inhibition by glyphosate. FEBS Lett. 1983. 154: 127-33
323. Cattani D, de Liz Oliveira Cavalli VL, Heinz Rieg CE, Domingues JT, Dal-Cim T, Tasca CI. Mechanisms underlying the neurotoxicity induced by glyphosate-based herbicide in immature rat hippocampus: Involvement of glutamate excitotoxicity. Toxicology. 2014. 320: 34-45
324. Cattani Dde Liz Oliveira Cavalli VLHeinz Rieg CEDomingues JTDal-Cim TTasca CILast accessed on 2014 Nov 22. Available from: http://wwwbfrbundde/en/the_bfr_has_finalised_its_draft_report_for_the_re_evaluation_of_glyphosate-188632html 2014 .
325. Mesnage RS, Bhat R, Gómez-López VM.editors. The Need for a Closer Look at Pesticide Toxicity during GMO Assessment. in Practical Food Safety: Contemporary Issues and Future Directions. Chichester, UK: John Wiley and Sons, Ltd; 2014. p.
326. . U.S.EPA. R.E.D. FACTS-Glyphosate. 1993. p.
327. Vandenberg LN, Colborn T, Hayes TB, Heindel JJ, Jacobs DR, Lee DH. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr Rev. 2012. 33: 378-455
328. Williams AL, Watson RE, Desesso JM. Developmental and reproductive outcomes in humans and animals after glyphosate exposure: A critical analysis. J Toxicol Environ Health B Crit Rev. 2012. 15: 39-96
329. Antoniou M, Habib ME, Howard CV, Jennings RC, Leifert C, Nodari RO. Teratogenic Effects of Glyphosate-Based Herbicides: Divergence of Regulatory Decisions from Scientific Evidence. J Environ Anal Toxicol. 2012. S4: 006-
330. Boocock MR, Coggins JR. Kinetics of 5-enolpyruvylshikimate-3-phosphate synthase inhibition by glyphosate. FEBS Lett. 1983. 154: 127-33
331. Cattani D, de Liz Oliveira Cavalli VL, Heinz Rieg CE, Domingues JT, Dal-Cim T, Tasca CI. Mechanisms underlying the neurotoxicity induced by glyphosate-based herbicide in immature rat hippocampus: Involvement of glutamate excitotoxicity. Toxicology. 2014. 320: 34-45
332. Cattani Dde Liz Oliveira Cavalli VLHeinz Rieg CEDomingues JTDal-Cim TTasca CILast accessed on 2014 Nov 22. Available from: http://wwwbfrbundde/en/the_bfr_has_finalised_its_draft_report_for_the_re_evaluation_of_glyphosate-188632html 2014 .
333. Mesnage RS, Bhat R, Gómez-López VM.editors. The Need for a Closer Look at Pesticide Toxicity during GMO Assessment. in Practical Food Safety: Contemporary Issues and Future Directions. Chichester, UK: John Wiley and Sons, Ltd; 2014. p.
334. . U.S.EPA. R.E.D. FACTS-Glyphosate. 1993. p.
335. Vandenberg LN, Colborn T, Hayes TB, Heindel JJ, Jacobs DR, Lee DH. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr Rev. 2012. 33: 378-455
336. Williams AL, Watson RE, Desesso JM. Developmental and reproductive outcomes in humans and animals after glyphosate exposure: A critical analysis. J Toxicol Environ Health B Crit Rev. 2012. 15: 39-96


Ian Brown
Posted October 15, 2016, 9:25 pm
Absolutely wonderful work, please never give up
Half of All Children Will Be Autistic by 2025, Warns Senior Research Scientist at MIT – Heatlh news and more
Posted November 15, 2016, 9:32 am
[…] Anthony Samsel and Stephanie Seneff. “Glyphosate, pathways to modern diseases III: Manganese, neurological diseases, and associated pathologies.” Surgical Neurology International 2015; 6:45. (Download) […]